





Citation: Mudzengi CP, Kativu S, Dahwa E, Poshiwa X, Murungweni C (2014) Effects of Dichrostachys cinerea (l.) Wight & Arn (Fabaceae) on herbaceous species in a semi-arid rangeland in Zimbabwe. Nature Conservation 7: 51–60. doi: 10.3897/natureconservation.7.5264
Anthropogenic alteration of an environment and other disturbance regimes may enable the expansion of some native species into new geographical areas, a phenomenon observed with Dichrostachys cinerea. Five D. cinerea invaded sites, each approximately one hectare in size were assessed for the effects of D. cinerea on native herbaceous species diversity, richness, basal cover, litter cover, top hamper and plant vigour. The same attributes were studied in five uninvaded sites adjacent to, and equal in size to each invaded site. Forty herbaceous species were identified in the area. There were significant differences (P < 0.05) noted in species richness, basal cover, litter cover, top hamper, plant vigour, and species diversities between invaded and uninvaded sites, with uninvaded sites recording higher values than invaded sites. Altitude, erosion and the edaphic variables pH, N, P and K, which were included as explanatory variables, also differed significantly (P<0.05) between invaded and uninvaded sites. Of the 30 D. cinerea invaded plots established for herbaceous species assessments, 26 were positively correlated with altitude, erosion, pH, P, N and K. It is imperative to find ways of managing D. cinerea in order to reduce its adverse effects on herbaceous species.
Dichrostachys cinerea, herbaceous species, invasion
Ecologically unsustainable anthropogenic activities such as agriculture, mining, and oil exploration, coupled with climate change and variability, facilitate plant species invasions. This is exacerbated by increases in the frequency and extent of natural disturbances such as droughts, hail storms, fire, insect outbreaks and disease in boreal forests which upset ecological and economic balances, perpetrating invasions (
Dichrostachys cinerea is a deciduous low thorny tree or shrub that produces bicoloured fragrant flowers. It is native to Africa and Asia, where it is common in the tropics of Africa, the Sahelian and Sudanian ecozones, and the South Arabian Peninsula (
Collection and grinding of Dichrostachys cinerea fruits have been suggested as a management tool to control its encroachment (
Rangelands on Gokomere Farm located 18 km north of Masvingo town, and covering approximately 5800 hectares of land were studied. The farm lies in Natural Region IV (NR IV) of the Zimbabwean ecological classification system (
A 2009 Google Earth Satellite Imagery of Gokomere Farm, followed by ground truthing was used to identify areas invaded by Dichrostachys cinerea Five Dichrostachys cinerea patches were chosen using the nearest neighbour-plus-one-method. Three transects measuring 100 m each were laid down, the first one passing through the centre of each patch, and each of the other two at equi-distances from the middle and boundary of each patch. On each transect, two 10 m × 10 m plots were systematically pegged on the ground. They were established at least 50 m from main roads and rivers to reduce road and river effects. Species presence/absence data and other herbaceous assessments were carried out in five 1 m × 1 m quadrats established at the four corners and centre of each 10 m × 10 m plot (
SPSS Version 13.0 (2004) was used for one way analyses of variance (ANOVA) of the herbaceous and soil variables. PAST was used to calculate diversity indices. The herbaceous variables that were analysed were basal cover, litter cover, plant vigour and top hamper. Altitude, erosion and the edaphic variables (pH, N, P and K) were included as explanatory variables. Multiple comparisons were done to test for significant differences among the plots. The relationships between the measured variables and the measured explanatory variables were explored using CANOCO for Windows (version 4). CANOCO was used to carry out Detrended Correspondence Analysis (DCA) and Redundancy Analysis (RCA) (
There was a total of 40 herbaceous species in the Dichrostachys cinerea invaded and uninvaded sites. Invaded sites had 26 species while uninvaded sites had 32. The most common species in the invaded sites were Digitaria penzii, Cynodon dactylon and Eragrostis trichophora while in the uninvaded sites they were Digitaria penzii, Eragrostis trichophora and Hyperthelia dissoluta (Table 1).
There were significant differences (F9, 50=9.375, P<0.05) in basal cover among invaded and uninvaded sites. Invaded sites did not differ significantly (F9, 50=2.750, P>0.05) in basal cover among themselves. However, the uninvaded sites showed significant differences (F9, 50=16.123, P<0.05). Litter cover had significant differences among invaded and uninvaded sites (F9, 50=6.024; P<0.05). Invaded sites were significantly different from each other (F9, 50=3.442, P<0.05), as were the uninvaded sites (F9, 50=6.537, P<0.05). Invaded sites recorded lower litter cover than uninvaded sites. Plant vigour in invaded sites was significantly different from that within uninvaded sites (F9, 50=5.796, P<0.05). Both the invaded sites and uninvaded sites showed significant differences in plant vigour (F9, 50=8.037, P<0.05 and F9, 50=4.605, P<0.05 respectively) among themselves. There were significant differences in top hamper among invaded and uninvaded sites (F9, 50=3.264, P<0.05). Invaded sites did not differ significantly (F9, 50=0.666, P>0.05) in top hamper. Uninvaded sites had significant differences (F9, 50=6.959, P<0.05). Species richness differed significantly (F9, 50=5.776, P<0.05) among invaded and uninvaded sites. Invaded sites were not significantly different (F9, 50=1.052, P>0.05) from each other in species richness. However, uninvaded sites had significant differences (F9, 50=7.113, P<0.05). Herbaceous species diversities also differed significantly (P<0.05) between invaded and uninvaded sites.
The pH values across invaded and uninvaded sites were significantly different (F9, 50=2.816, P<0.05). Invaded sites had significant differences for pH among themselves (F9, 50=4.738, P<0.05) unlike uninvaded sites (F9, 50=1.719, P>0.05). There were significant differences (F9, 50=70.903, P<0.05) for K between invaded and uninvaded sites. Both, invaded and uninvaded sites were significantly different from each other (F9, 50=100.952, P<0.05; F9, 50=4.535, P<0.05). The recorded P values showed significant differences (F9, 50=66.916, P<0.05) between invaded and uninvaded values. Both invaded and uninvaded sites showed significant differences among themselves (F9, 50=51.018, P<0.05 and F9, 50=24.872, P<0.05 respectively). There were significant differences (F9, 50=3.980, P<0.05) for the recorded N values between the invaded and uninvaded sites. Uninvaded sites significantly differed from each other (F9, 50=5.039, P<0.05) while invaded sites were not significantly different (F9, 50=2.735, P>0.05). There were significant differences (F9, 50=6.193, P<0.05) in altitude between the invaded and uninvaded sites. Invaded sites were significantly different (F9, 50=5.692, P<0.05) from each other. Uninvaded sites also significantly differed (F9, 50=7.637, P<0.05) from each other. Erosion significantly differed (F9, 50=3.303, P<0.05) between invaded and uninvaded sites. Invaded sites had significant differences for erosion among themselves (F9, 50=4.420, P<0.05), and so did uninvaded sites (F9, 50=3.182, P<0.05).
Of the 30 Dichrostachys cinerea invaded plots (1–30) assessed for herbaceous species attributes, 26 were correlated with altitude, erosion, pH, P, N and K (Figure 1). Twenty five of the uninvaded plots (31–60), were negatively correlated with the measured environment variables. Invaded plots classified with these uninvaded sites were 6, 22, 23 and 25. They had low K values characteristic of the uninvaded sites studied, with the exception of sites 4 and 6. They all had lower N than the invaded plots (0.5–2.1 ppm compared to 3.3–5 ppm). There was no clear pattern for pH.
The species that were negatively correlated with these variables were Aristida spp., Pogonarthria squarrosa, Rhynchelytrum repens, Panicum spp. and Digitaria penzii. K was positively correlated with Urochloa mozambicensis, Heteropogon contortus, Acanthospermum hispidum etc. It was negatively correlated with Richadia scabra, Setaria palidifusca and Hyperthelia dissoluta (Fig. 2).
We show that pH, erosion, P, altitude and N influenced the abundance of species such as Hibiscus spp, Sida alba and Cynodon dactylon.
Herbaceous species variables among the invaded and uninvaded sites.
| Parameter | Invaded Site | Uninvaded Site |
|---|---|---|
| Basal cover | 3.7a | 4.5b |
| Litter cover | 1.5a | 2.1b |
| Species richness | 2.9a | 3.5b |
| Plant vigour | 2.9a | 3.4b |
| Top hamper | 0.8a | 1.1b |
| Shannon_H index | 2.58a | 2.85b |
| Simpson_1-D index | 0.92a | 0.94b |
Means in rows with different superscripts are significantly different (P<0.05)
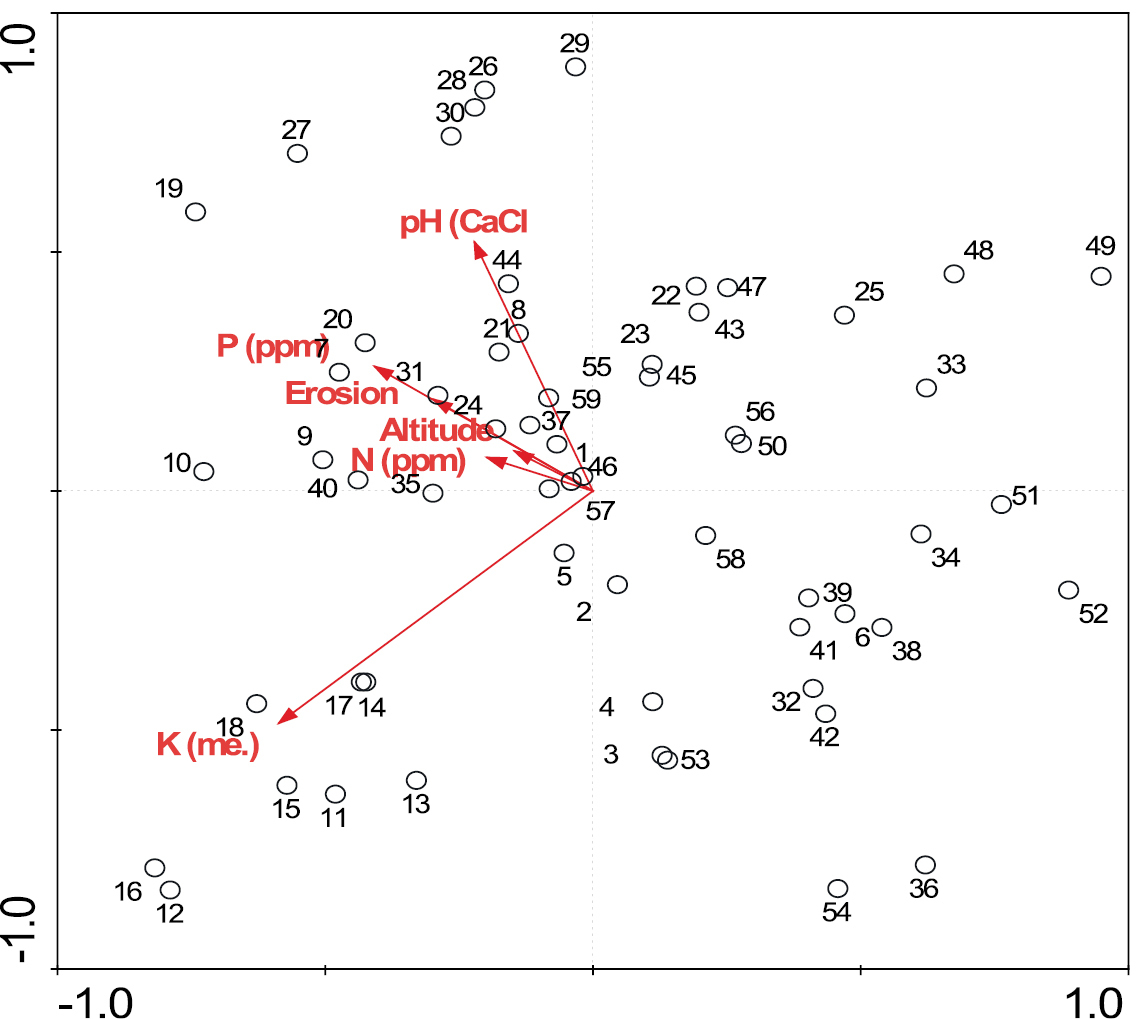
A redundancy analysis plot based on herbaceous species presence absence data showing relationship between the environmental variables and the plots.

A redundancy analysis plot based on herbaceous species presence absence data showing relationship between the environmental variables and the species.
Herbaceous variables
Dichrostachys cinerea invasion has caused declines in herbaceous species richness. Competitive interactions are critical between woody plants and grass dominated layers in arid to semi-arid areas (
In a study by
In the present study there were significant differences in environmental variables between invaded and uninvaded sites. Invaded sites had higher soil N, K and P than uninvaded sites. Biological invaders change ecosystems as they differ from native species in resource acquisition and/or resource use efficiency. They may also alter the trophic structure of the area invaded, or the disturbance frequency and/or intensity (
Eragrostis spp., Hibiscus spp., Sida alba and Urochloa mozambicensis were found in the Dichrostachys cinerea invaded sites. These grasses are associated with disturbed land (
The herbaceous species that were negatively correlated with the measured environmental variables were Aristida spp., Philodendron maximum, Pogonarthria squarrosa, Digitaria penzii, Hyparrhenia filipendula, Hyperthelia dissoluta and Rhynchelytrum repens among other species. Philodendron maximum grows best in moist, well-drained soils just like Rhynchelytrum repens, although some of its varieties are tolerant of lower fertility and poorer drainage (
Dichrostachys cinerea is a native invasive woody shrub or tree. Here, it adversely affected native herbaceous species plant vigour, basal cover and species richness. Reduced litter cover and top hamper were also observed in invaded sites. These observations can be attributed to the fast growth, propagation and propagule pressure that characterize Dichrostachys cinerea, giving it a competitive advantage with respect to acquisition of light, nutrients and other resources. Therefore, these adverse effects on herbaceous species may, in the long term, reduce the carrying capacity of rangelands, making them even more susceptible to alien invaders. However, a long-term study may provide more information on the biology and invasive capacity of the species and on its impact on other vegetation and animal species. There is also need to find environmentally friendly and effective methods of controlling Dichrostachys cinerea inorder to prevent its spread, hence adverse effects on herbaceous species.