Research Article |
|
Corresponding author: Abdul Rashid Jamnia ( a.r.jamnia@gmail.com ) Academic editor: Maurizio Pinna
© 2018 Abdul Rashid Jamnia, Ahmad Ali Keikha, Mahmoud Ahmadpour, Abdoul Ahad Cissé, Mohammad Rokouei.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Jamnia AR, Keikha AA, Ahmadpour M, Cissé AA, Rokouei M (2018) Applying bayesian population assessment models to artisanal, multispecies fisheries in the Northern Mokran Sea, Iran. Nature Conservation 28: 61-89. https://doi.org/10.3897/natureconservation.28.25212
|
Abstract
Small-scale fisheries substantially contribute to the reduction of poverty, local economies and food safety in many countries. However, limited and low-quality catches and effort data for small-scale fisheries complicate the stock assessment and management. Bayesian modelling has been advocated when assessing fisheries with limited data. Specifically, Bayesian models can incorporate information of the multiple sources, improve precision in the stock assessments and provide specific levels of uncertainty for estimating the relevant parameters. In this study, therefore, the state-space Bayesian generalised surplus production models will be used in order to estimate the stock status of fourteen Demersal fish species targeted by small-scale fisheries in Sistan and Baluchestan, Iran. The model was estimated using Markov chain Monte Carlo (MCMC) and Gibbs Sampling. Model parameter estimates were evaluated by the formal convergence and stationarity diagnostic tests, indicating convergence and accuracy. They were also aligned with existing parameter estimates for fourteen species of the other locations. This suggests model reliability and demonstrates the utility of Bayesian models. According to estimated fisheries’ management reference points, all assessed fish stocks appear to be overfished. Overfishing considered, the current fisheries management strategies for the small-scale fisheries may need some adjustments to warrant the long-term viability of the fisheries.
Keywords
Small-scale Demersal fisheries, Bayesian modelling, Generalised surplus production
Introduction
Human dependency on maritime and coastal resources is increasing (
Increased over-exploitation of fishery and habitat destruction threaten the coastal and maritime resources. Small-scale fisheries often have limited and low-quality catch and effort data that complicates stock assessment and management. Globally, for example, only 10% to 50% of fish stocks in more developed countries and 5% to 20% of fish stocks in less-developed countries have been scientifically assessed due to limited data (
Based on the various works (
Mostly, due to insufficient information about the time series of biological and management reference points of fish stocks, the scientific precise stock assessments have not been undertaken for the majority of fish species in Iran, especially in the southern coastal areas (the current study area). Therefore, due to the lack of information toward fisheries management reference points, most fisheries planning has not had any special effects on the sustainability of fish stocks reserves.
So far, there has been a lot of scientific motivation for assessing fish stocks in Iran, but it has not been implemented for two reasons; one the lack of sufficient information used in scientific stock assessment methods and second the complexity and time-consuming of stock assessment methods in the limited-data situations.
Bayesian modelling has been advocated to assess fisheries with limited data. Specifically, Bayesian models can incorporate information of the multiple sources such as academic literature, empirical research, biological theory and specialist judgement. This characteristic of the Bayesian models improves precision in the stock assessments and provides the specific levels of uncertainty for estimating the parameters (
Therefore, in the current study, because of the limited data, the state-space Bayesian generalised surplus production models used to estimate the stocks status of the fourteen demersal fish species targeted by small-scale fisheries in Sistan and Baluchestan (A coastal province south-east of Iran). This could provide scientific knowledge for the fisheries management and contribute to the researchers applying and improving the results of the current study in order to achieve the global environmental sustainability and marine ecology.
Hence, in the current study, based on the above-mentioned stock assessment Bayesian approach, the management reference points provided for the fourteen fish species, including biomass, harvest rate and stock status, the implication of these for the sustainable management of the small-scale fisheries was discussed. Moreover, the estimates of biological parameters were compared to the previous findings of the fourteen species.
Materials, methods and data selection
Study area and data source
The fisheries examined in this study are located in the Sistan and Baluchestan Province (SBP) situated at the northern end of Mokran Sea (Gulf of Oman) in Iran (

Based on the local reportage of PFDSB and IFO, the fishery is an important source of income, cultural heritage and recreation in the area. In addition, it forms the largest employment with over 24,500 locals involved in fishing industries permanently or seasonally. The fishing system of SBP consists of inshore fleets, with 60% (1430) of vessels registered as weighing at least three gross tonnes, as most of which are primarily made of fibreglass. The fishing activities are mainly seasonal and fishermen change their fishing gear (gill nets, hand-lines/hook-and-lines and traps) and strategies based on water levels, habitats, migration patterns and species targeted. However, the majority of vessels that fish annually use gill nets. The total catches of the fourteen species examined in this study from 21 March 2015 to 18 March 2016 were 35,937 metric tonnes. This catch comprises 49% of the demersal fish catch; 31% of all fish species caught and 187 million US$ of economic value which represents an important resource for the fishery.
Nine years (21 March 2006 to 18 March 2015) of fisheries-dependent commercial landings and effort data for the fourteen demersal fish species listed earlier were acquired from Iran’s National Fishery Data Collection and Reporting System Unit and also reports from the Provincial Fisheries Department. In addition, the used time series of catch and standardised CPUE values as relative abundance indices are considerable as the supplementary data. The nominal CPUE indices that derived from commercial fisheries’ logbooks are affected by some variables such as spatiotemporal and environmental factors. The considered standardised CPUE indices are reliable abundance indices which allow the implementation of the conservation and management measures and have been obtained by the most common and competent statistical approach in the domain of fisheries’ researches, such as generalised additive models (GAMs), that are used for standardising catch and effort data.
Modelling methods
Biomass dynamic models are popularly used for stock assessment when only catch and effort time series data are accessible (
The biomass dynamics model of the equation discrete time form is as follows (
Bt = Bt–1+ h (Bt–1) – Ct–1 (1)
In Equation 1, Bt–1, Ct– and h (Bt–1) denote biomass and catch for year t–1 and the surplus production function, respectively.
Bayesian state-space surplus production model
Bayesian state-space models consist of three levels (
(I) a process equation which depicts the time dynamics of a stochastic process as a function of time-invariant hyper-parameters. (II) an observation equation based on population-specific inspection data that are a function of the unobserved state process (
Process equation
With regards to the Equation 1, the process equation describes the surplus production function in a generalised surplus production model (GSPM) (
h (Bt–1) = r Bt–1(1 – (Bt–1 / K)z), r > 0, K > 0, z > 0 (2)
where r is the intrinsic population growth rate; K is the carrying capacity of the population and z is the shape parameter of the production model that determines at which B/K ratio maximum surplus production was attained and commonly noted as equivalent biomass and at which the maximum sustainable yield (MSY) was attained (BMSY). If the shape parameter was less than unity (0 < z < 1), then surplus production would increase (to the peak point) when the biomass was below K/2 (a left-skewed production curve). If the shape parameter was greater than unity (z > 1), then biomass production would increase (to the peak point) when the biomass was more than K/2 (a right-skewed production curve). If the shape parameter was identical to unity (z = 1), the production model would reduce to the Schafer form, attaining MSY when biomass was equal to K/2. If z approached zero (z → 0), the production model would reduce to the Fox model that results into maximum surplus production at ~0.37K.
Replacing Equation 2 in Equation 1 and multiplying the right hand side of the resultant equation with ut yields the stochastic form of the biomass dynamic model with generalised surplus production (GSP) (
Bt = (Bt–1 + rtBt–1(1 – (Bt–1 / K)z) – Ct–1)ut (3)
Where ut is process noise – supposed to be independent and log-normally distributed; specifically ut = eεt where εt ~ N[0, σ2], i.e. εt is i.i.d. normal with mean zero and variance σ2.
Equation 3 was re-parameterised using relative biomass (P = Bt / K) to diminish parameter confounding such as that between biomass and K that could result in related priors (
Pt = (Pt–1 + rtPt–1(1 – (Pt–1)z) – Ct–1 / K)ut (4)
Observation equation
According to regular assumptions, CPUE values are relative abundance indices proportional to the biomass. The observation equation relates the unobserved states Bt to the relative abundance indices It (
It = qKPt υt (5)
where It is the relative biomass index; q is the “catchability” coefficient that indicates the effectiveness of each unit of fishing effort and υt is the observation error entered as an independent and log-normally distributed random variable. Specifically, υt = eηt where ηt ~ N[0, τ2], i.e. is ηt i.i.d. normal with mean zero and variance τ2.
Parameter prior distribution layer
An advantage of the Bayesian models is its ability to use the prior distributions based upon the existing knowledge to set plausible values for model parameters (
The reason for choosing the priors was based on the following rationale. First, based on the expert consultations of IFO and PFDSB and the available information from
Summary of the Prior distribution functions used for some parameters of all bayesian state-space GSPM Fourteen specified high-commercial demersal fish species.
| Species | Parameter | ||
| Intrinsic growth rate, r | Carrying capacity, K | Initial condition, P[1] | |
| Elutheronema tetradactylum | r ~ lognormal(-1.1139,4.4814) | K ~ uniform(604,30200) | pi ~ uniform(0.01,0.135)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Lethrinus microdon | r ~ lognormal(-0.2326,4.4814) | K ~ uniform(1089,54450) | pi ~ uniform(0.0132,0.125)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Lutjanus johni | r ~ lognormal(-1.4741,4.4814)) | K ~ uniform(1180,59000) | pi ~ uniform(0.019,0.116)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Lutjanus malabaricus | r ~ lognormal(-1.2447,4.4814) | K ~ uniform(704,35200) | pi ~ uniform(0.012,0.14)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Otolithes ruber | r ~ lognormal(-1.9888,4.4814) | K ~ uniform(4187,209350) | pi ~ uniform(0.01,0.135)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Pampus argenteus | r ~ lognormal(-0.5547,4.4814) | K ~ uniform(569,28450) | pi ~ uniform(0.014,0.132)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Parastromateus niger | r ~ lognormal(-1.8574,4.4814) | K ~ uniform(6456,322800) | pi~uniform(0.01,0.121)pm=log(pi)P[1]~lognormal(pm, 1/σ2) |
| Platycephalus indicus | r ~ lognormal(-2.0425,4.4814) | K ~ uniform(1076,53800) | pi ~ uniform(0.0126,0.113)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Pomadasys kaakan | r ~ lognormal(0.0615,4.4814) | K ~ uniform(2992,149600) | pi ~ uniform(0.01,0.114)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Protonibea diacanthus | r ~ lognormal(-1.0531,4.4814) | K ~ uniform(4429,221450) | pi ~ uniform(0.0171,0.1587)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Saurida tumbil | r ~ lognormal(-0.9886,4.4814) | K ~ uniform(2170,108500) | pi ~ uniform(0.0154,0.1286)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Scomberoides commersonnianus | r ~ lognormal(-1.5099,4.4814) | K ~ uniform(11001,55005) | pi ~ uniform(0.0194,0.105)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Rhabdosargus sarba | r ~ lognormal(-1.0634,4.4814) | K ~ uniform(1746,87300) | pi ~ uniform(0.0164,0.1613)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
| Trachinotus mookalee | r ~ lognormal(-1.7514,4.4814) | K ~ uniform(712,35600) | pi ~ uniform(0.0163,0.12)pm = log(pi)P[1] ~ lognormal(pm, 1/σ2) |
Based on
Fisheries management reference points
The major fisheries reference points for the GSPM (
The stock biomass at which the maximum sustainable yield (MSY) was attained (BMSY) is given as Equation 6:
BMSY = K (z + 1)(-1/z) (6)
whilst, FMSY is the fishing mortality corresponding to MSY and is described as Equation 7:
FMSY = K (z + 1)(-1/z) (7)
The related value of MSY is calculated as Equation 8:
MSY = BMSY FMSY = K (z + 1)(-1/z).r (z + 1)(-1/z) = Kr (z + 1)(-2/z) (8)
Finally, the relative fishing mortality rate FS and relative biomass BS are assessed by FS = Ft / FMSY and BS = Bt / BMSY , respectively.
Model fitting
The Bayesian state-space combines the joint prior distributions of all parameters and unobservable conditions with the likelihood functions of the observations (
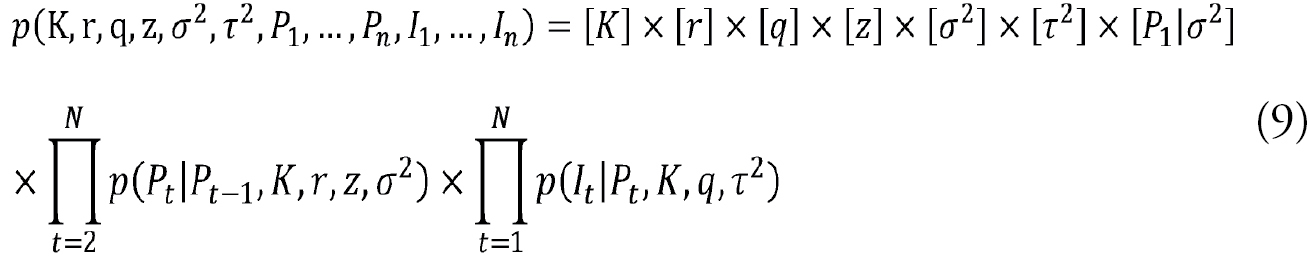
with square brackets indicating densities and N referring to the number of samples. Supplementary information on the above-mentioned general factorisation of Bayesian model (Eq. 9) is available in
The Markov chain Monte Carlo (MCMC) algorithm using Gibbs sampling was used to explore the joint posterior distributions of the parameters and un-observable states. OpenBugs software (v3.2.3) (
Results and discussion
In line with the aims of the current research, the results of Bayesian state-space GSPM from MCMC simulations are briefly presented in Tables


Convergence to posterior distribution
Table
Convergence and stationarity diagnostics of MCMC algorithm for bayesian state-space gspm fourteen specified high-commercial demersal fish species.
| Species | Geweke’s Z-score | Gelman-Rubin | Heidelberger-Welch’s p-value | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chain 1 | Chain 2 | Chain 3 | Potential scale reduction factor (Ȓ) | Chain 1 | Chain 2 | Chain 3 | |||||||||||||||
| Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Chain 1 | Chain 2 | Chain 3 | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | |
| Elutheronema tetradactylum | -1.73 | 1.48 | 0.228 | -1.86 | 1.62 | -0.065 | -1.72 | 1.452 | 0.135 | 1 | 1 | 1 | 0.006 | 0.9 | 0.383 | 0.05 | 0.9 | 0.546 | 0.027 | 0.97 | 0.4 |
| Lethrinus microdon | -1.7 | 1.36 | 0.245 | -1.64 | 1.08 | 0.09 | -1.66 | 1.61 | 0.174 | 1 | 1 | 1 | 0.014 | 0.9 | 0.496 | 0.054 | 0.96 | 0.568 | 0.053 | 0.98 | 0.4 |
| Lutjanus johni | -1.36 | 1.13 | 0.227 | -1.14 | 1.48 | -0.129 | -1.7 | 1.36 | 0.22 | 1 | 1 | 1 | 0.004 | 0.95 | 0.447 | 0.058 | 0.91 | 0.513 | 0.052 | 0.99 | 0.68 |
| Lutjanus malabaricus | -1.33 | 1.261 | 0.06 | -1.6 | 1.612 | -0.05 | -1.02 | 1.51 | 0.407 | 1 | 1 | 1 | 0.03 | 0.99 | 0.67 | 0.05 | 0.99 | 0.486 | 7E-6 | 0.99 | 0.71 |
| Otolithes ruber | -1.79 | 1.365 | 0.068 | -1.55 | 1.52 | 0.252 | -1.77 | 1.4 | 0.264 | 1 | 1 | 1 | 0.007 | 0.99 | 0.619 | 0.007 | 0.99 | 0.568 | 0.07 | 0.98 | 0.477 |
| Pampus argenteus | -1.83 | 1.9 | 0.195 | -1.01 | 1.03 | 0.188 | -1.47 | 1.43 | 0.345 | 1 | 1 | 1 | 0.01 | 0.98 | 0.649 | 0.05 | 0.99 | 0.512 | 0.056 | 0.99 | 0.677 |
| Parastromateus niger | -1.06 | 1.98 | 0.181 | -1.47 | 1.37 | 0.11 | -1.45 | 1.84 | 0.02 | 1 | 1 | 1 | 0.05 | 0.98 | 0.391 | 0.05 | 0.98 | 0.44 | 0.01 | 0.99 | 0.412 |
| Platycephalus indicus | -1.25 | 1.56 | 0.11 | -1.15 | 1.9 | 0.11 | -1.35 | 1.61 | 0.232 | 1 | 1 | 1 | 0.05 | 0.99 | 0.39 | 0.06 | 0.99 | 0.44 | 0.006 | 0.99 | 0.61 |
| Pomadasys kaakan | -1.11 | 1.91 | 0.018 | -1.63 | 1.3 | 0.251 | -1.88 | 1.06 | 0.268 | 1 | 1 | 1 | 0.01 | 0.99 | 0.7 | 0.07 | 0.95 | 0.49 | 0.002 | 0.99 | 0.62 |
| Protonibea diacanthus | -1.27 | 1.03 | 0.226 | -1.1 | 1.35 | 0.33 | -1.14 | 1.8 | -0.02 | 1 | 1 | 1 | 0.007 | 0.98 | 0.623 | 0.05 | 0.99 | 0.476 | 0.008 | 0.99 | 0.592 |
| Saurida tumbil | -1.18 | 1.11 | 0.035 | -1.18 | 1.78 | 0.007 | -1.08 | 1.24 | 0.176 | 1 | 1 | 1 | 0.05 | 0.99 | 0.49 | 0.07 | 0.97 | 0.34 | 0.06 | 0.98 | 0.57 |
| Scomberoides commersonnianus | -1.19 | 1.14 | 0.19 | -1.13 | 1.69 | 0.126 | -1.04 | 1.53 | 0.204 | 1 | 1 | 1 | 0.05 | 0.9 | 0.5 | 0.007 | 0.99 | 0.297 | 0.05 | 0.95 | 0.48 |
| Rhabdosargus sarba | -1.25 | 1.51 | 0.13 | -1.23 | 1.21 | 0.12 | -1.04 | 1.07 | 0.048 | 1 | 1 | 1 | 0.05 | 0.9 | 0.5 | 0.05 | 0.9 | 0.5 | 0.02 | 0.98 | 0.61 |
| Trachinotus mookalee | -1.89 | 1.62 | 0.024 | -1.2 | 1.72 | 0.242 | -1.55 | 1.25 | 0.093 | 1 | 1 | 1 | 0.05 | 0.98 | 0.36 | 0.05 | 0.99 | 0.417 | 0.017 | 0.96 | 0.5 |
In the first place, the Geweke diagnostic test was separately applied to verify convergence of the mean of each parameter obtained from the sampled values related to each single chain. In the following, the derived Z-score indicates convergence if its values be less than 2 at absolute value. Thus, as shown in Table
Estimates of model parameters
Summary of the posterior descriptive statistics is presented in Table
A summary of the posterior descriptive statistics for bayesian state-space GSPM parameters of fourteen specified high-commercial demersal fish species.
| Parameter | Species | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elutheronema tetradactylum | Lethrinus microdon | Lutjanus johni | Lutjanus malabaricus | Otolithes ruber | Pampus argenteus | Parastromateus niger | Platycephalus indicus | Pomadasys kaakan | Protonibea diacanthus | Saurida tumbil | Scomberoides commersonnianus | Rhabdosargus sarba | Trachinotus mookalee | ||
| r | Mean | 0.329 | 0.727 | 0.248 | 0.298 | 0.152 | 0.612 | 0.34 | 0.145 | 1.173 | 0.474 | 0.411 | 0.247 | 0.378 | 0.175 |
| S.D. | 0.141 | 0.286 | 0.113 | 0.134 | 0.074 | 0.279 | 0.161 | 0.073 | 0.567 | 0.328 | 0.201 | 0.124 | 0.175 | 0.079 | |
| MC error | 0.001 | 0.004 | 0.001 | 0.001 | 9.8E-4 | 0.002 | 0.001 | 8.3E-4 | 0.004 | 0.009 | 0.002 | 0.001 | 0.002 | 7.6E-4 | |
| Perc. 2.5% | 0.126 | 0.295 | 0.09 | 0.111 | 0.054 | 0.223 | 0.124 | 0.051 | 0.42 | 0.142 | 0.146 | 0.087 | 0.136 | 0.066 | |
| Median | 0.304 | 0.688 | 0.223 | 0.273 | 0.137 | 0.558 | 0.306 | 0.13 | 1.057 | 0.377 | 0.369 | 0.221 | 0.343 | 0.16 | |
| Perc. 97.5% | 0.668 | 1.393 | 0.529 | 0.627 | 0.336 | 1.295 | 0.748 | 0.332 | 2.598 | 1.314 | 0.92 | 0.561 | 0.806 | 0.367 | |
| K | Mean | 1058 | 1530 | 1995 | 1110 | 5175 | 1409 | 12080 | 1609 | 4015 | 6641 | 2580 | 11630 | 2146 | 1121 |
| S.D. | 365.6 | 370 | 623.6 | 353 | 908.7 | 553.5 | 5477 | 445.7 | 902.6 | 1876 | 390.6 | 616.1 | 365.3 | 336 | |
| MC error | 2.726 | 3.717 | 6.953 | 2.64 | 18.11 | 5.416 | 51.99 | 4.183 | 6.027 | 28.51 | 4.339 | 6.865 | 4.086 | 2.851 | |
| Perc. 2.5% | 621.5 | 1104 | 1214 | 718.1 | 4215 | 634.5 | 6602 | 1095 | 3022 | 4504 | 2182 | 11020 | 1758 | 727.4 | |
| Median | 975.7 | 1438 | 1865 | 1017 | 4918 | 1318 | 10450 | 1498 | 3771 | 6149 | 2466 | 11450 | 2042 | 1041 | |
| Perc. 97.5% | 1975 | 2463 | 3526 | 2018 | 7565 | 2736 | 26630 | 2734 | 6334 | 11510 | 3607 | 13270 | 3109 | 1971 | |
| q | Mean | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 5.5E-4 | 0.001 | 0.001 | 0.001 | 9.5E-4 | 4.3E-4 | 0.001 | 0.001 |
| S.D. | 5E-4 | 3E-4 | 4.4E-4 | 4.2E-4 | 1.5E-4 | 7.2E-4 | 1.9E-4 | 3.4E-4 | 2.5E-4 | 3.2E-4 | 1.3E-4 | 3.2E-5 | 1.6E-4 | 4E-4 | |
| MC error | 4E-6 | 3E-6 | 4.5E-6 | 3E-6 | 3E-6 | 6.5E-6 | 1.7E-6 | 3E-6 | 1.6E-6 | 4.9E-6 | 1.5E-6 | 5E-7 | 1.8E-6 | 3.2E-6 | |
| Perc. 2.5% | 8E-4 | 8.8E-4 | 8.4E-4 | 8.3E-4 | 6.9E-4 | 8.5E-4 | 2E-4 | 8E-4 | 7.7E-4 | 6.8E-4 | 6.6E-4 | 3.6E-4 | 6.9E-4 | 8.4E-4 | |
| Median | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 5.5E-4 | 0.001 | 0.001 | 0.001 | 9.5E-4 | 4.3E-4 | 0.001 | 0.001 | |
| Perc. 97.5% | 0.002 | 0.002 | 0.002 | 0.002 | 0.001 | 0.003 | 9.2E-4 | 0.002 | 0.001 | 0.001 | 0.001 | 4.9E-4 | 0.001 | 0.002 | |
| z | Mean | 0.953 | 0.89 | 0.995 | 0.96 | 0.993 | 0.987 | 0.983 | 1.001 | 0.996 | 1.047 | 1.006 | 0.982 | 0.993 | 0.965 |
| S.D. | 0.708 | 0.661 | 0.709 | 0.688 | 0.695 | 0.712 | 0.698 | 0.704 | 0.699 | 0.72 | 0.73 | 0.691 | 0.701 | 0.711 | |
| MC error | 0.006 | 0.008 | 0.008 | 0.005 | 0.012 | 0.006 | 0.005 | 0.005 | 0.005 | 0.01 | 0.011 | 0.012 | 0.007 | 0.005 | |
| Perc. 2.5% | 0.112 | 0.108 | 0.119 | 0.117 | 0.122 | 0.118 | 0.121 | 0.12 | 0.118 | 0.131 | 0.118 | 0.12 | 0.117 | 0.109 | |
| Median | 0.779 | 0.723 | 0.829 | 0.795 | 0.84 | 0.821 | 0.82 | 0.842 | 0.838 | 0.89 | 0.836 | 0.824 | 0.832 | 0.792 | |
| Perc. 97.5% | 2.736 | 2.604 | 2.808 | 2.707 | 2.728 | 2.776 | 2.766 | 2.774 | 2.752 | 2.869 | 2.824 | 2.726 | 2.765 | 2.803 | |
| σ2 | Mean | 0.007 | 0.007 | 0.008 | 0.007 | 0.007 | 0.007 | 2.5E-4 | 0.007 | 0.007 | 0.008 | 0.008 | 0.008 | 0.008 | 0.007 |
| S.D. | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 1.4E-4 | 0.002 | 0.002 | 0.003 | 0.002 | 0.003 | 0.003 | 0.002 | |
| MC error | 1.2E-5 | 1.5E-5 | 1.6E-5 | 1E-5 | 1.3E-5 | 1.4E-5 | 8.7E-7 | 1.6E-5 | 1.4E-5 | 2.3E-5 | 1.5E-5 | 1.8E-5 | 1.8E-5 | 1.2E-5 | |
| Perc. 2.5% | 0.004 | 0.004 | 0.004 | 0.004 | 0.004 | 0.004 | 9.8E-5 | 0.004 | 0.004 | 0.004 | 0.004 | 0.004 | 0.004 | 0.004 | |
| Median | 0.006 | 0.007 | 0.008 | 0.006 | 0.007 | 0.007 | 2E-5 | 0.007 | 0.007 | 0.008 | 0.007 | 0.008 | 0.007 | 0.007 | |
| Perc. 97.5% | 0.012 | 0.014 | 0.015 | 0.012 | 0.012 | 0.139 | 6.3E-4 | 0.014 | 0.013 | 0.017 | 0.014 | 0.016 | 0.015 | 0.012 | |
| τ2 | Mean | 0.358 | 0.071 | 0.021 | 0.459 | 0.012 | 0.079 | 0.121 | 0.068 | 0.082 | 0.027 | 0.056 | 0.028 | 0.046 | 0.145 |
| S.D. | 0.105 | 0.023 | 0.007 | 0.134 | 0.004 | 0.025 | 0.045 | 0.022 | 0.026 | 0.01 | 0.019 | 0.011 | 0.016 | 0.044 | |
| MC error | 6E-4 | 1.3E-4 | 4.8E-5 | 8E-4 | 2.5E-5 | 1.3E-4 | 3.2E-4 | 1.3E-4 | 1.4E-4 | 1E-4 | 1E-4 | 7.6E-5 | 9.9E-5 | 2.4E-4 | |
| Perc. 2.5% | 0.206 | 0.037 | 0.009 | 0.266 | 0.006 | 0.042 | 0.06 | 0.035 | 0.044 | 0.01 | 0.028 | 0.012 | 0.022 | 0.08 | |
| Median | 0.341 | 0.068 | 0.02 | 0.437 | 0.012 | 0.075 | 0.112 | 0.064 | 0.078 | 0.026 | 0.053 | 0.026 | 0.044 | 0.138 | |
| Perc. 97.5% | 0.613 | 0.127 | 0.04 | 0.785 | 0.023 | 0.139 | 0.236 | 0.122 | 0.146 | 0.053 | 0.102 | 0.055 | 0.085 | 0.252 | |
Since the intrinsic growth rate r shows the relationship between size and age, it is an important factor in life history theory (
According to the results in Table
A summary of model selection information between Generalised Surplus Production Model (GSPM) and Classic Schaefer (logistic) Surplus Production Model (SSPM) based on predictive performance using Deviance Information Criterion (DIC) for fourteen studied demersal fish species.
| Species | Models | |
|---|---|---|
| GSPM | SSPM | |
| Total DIC | Total DIC | |
| Elutheronema tetradactylum | 1.52 | 3.07 |
| Lethrinus microdon | 10.48 | 12.38 |
| Lutjanus johni | 22.33 | 25.59 |
| Lutjanus malabaricus | 2.352 | 3.87 |
| Otolithes ruber | 20.73 | 25.47 |
| Pampus argenteus | 7.267 | 9.49 |
| Parastromateus niger | 6.68 | 8.38 |
| Platycephalus indicus | 11.24 | 13.14 |
| Pomadasys kaakan | 7.14 | 8.96 |
| Protonibea diacanthus | 2.65 | 6.16 |
| Saurida tumbil | 11.82 | 13.92 |
| Scomberoides commersonnianus | 8.68 | 10.98 |
| Rhabdosargus sarba | 15.97 | 18.52 |
| Trachinotus mookalee | 8.02 | 9.68 |
The results of Table
The marginal posterior means of the catchability coefficient (q) for Elutheronema tetradactylum was within the 95% credibility intervals (0.0008, 0.002) and indicates that, for the given available data information (prior distribution), the true value of (q) falls within (Crl) with 95% probability. Similarly, for all other thirteen fish species, the marginal posterior means of (q) were within the 95% credibility intervals of the posterior predictive distributions and it can be concluded that for the given available data information (prior distribution), the true value of (q) falls within (Crl) with 95% probability for them. Furthermore, the posterior means and medians of the catchability coefficient (q) with 95% confidence interval for all the studied fish species were equal which shows that their posterior distributions were symmetric.
The marginal posterior means of the process noise variances, (σ2) for Elutheronema tetradactylum was within the 95% credibility intervals (0.004, 0.012) and indicates that, for the given available data information (prior distribution), the true value of (σ2) falls within (Crl) with 95% probability. Similarly, also, for all other thirteen fish species, the marginal posterior means of (σ2) were within the 95% credibility intervals of the posterior predictive distributions and it can be concluded that, for the given available data information (prior distribution), the true value of (σ2) falls within (Crl) with 95% probability for them. In addition, for the process noise variances of Bayesian state-space GSPM of Lutjanus malabaricus, Parastromateus niger, Saurida tumbil and Rhabdosargus sarba with 95% confidence interval, the posterior means were generally higher than the medians because their posterior distributions were right skewed. However, for the other 10 fish species, the posterior means and medians of the process noise variances Ó2 were equal which shows that their posterior distributions were symmetric.
The marginal posterior means of the observation noise variances, (τ2) for Elutheronema tetradactylum within 95% credibility intervals (0.206, 0.613), indicates that for the given available data information (prior distribution), the true value of (τ2) falls within (Crl) 95% probability. Similarly, for all other thirteen fish species, the marginal posterior means of (τ2) were within 95% credibility intervals of the posterior predictive distributions and it can be concluded that, for the given available data information (prior distribution), the true value of (τ2) falls within (Crl) with 95% probability for them. In addition, the means and medians marginal posterior of observation noise variances (τ2) of Bayesian state-space GSPM, except for Otolithes ruber, were dissimilar for the other species. For the above-considered fish species, the equality of the posterior mean and median of observation noise variances (τ2) indicates that their posterior distributions were symmetric. However, for all the other remaining 13 fish species, due to the right skewed of their posterior distributions, the means were generally higher than the medians.
Estimates of reference points
A summary of the results of fisheries management reference points derived from Bayesian state-space GSPM is graphically presented for all fourteen specified high-commercial demersal fish species through the stock status plots (i.e. Kobe plots or phase diagrams) in Figure
The Kobe plot characterises, relative biomass (BS = B / BMSY) and relative fishing mortality rate (FS = F / FMSY) in a graph which provides four different quadrants, each indicating a different population status. The red region is kept for the worst case in which the stock is excessively overfished (B / BMSY < 1) and, at the same time, the overfishing is at a high rate (F / FMSY > 1). The green zone denotes a situation where no overfishing is happening (F / FMSY < 1) and where the stock is not overfished (B / BMSY > 1), so it is the best condition for the stock. The orange quadrant presents a situation where overfishing is occurring (F / FMSY > 1), while the stock is not overfished (B / BMSY > 1), so a decrease in fishing intensity would bring it back to the ideal green condition. The yellow subdivision shows that the stock has been overfished (B / BMSY < 1), while the overfishing has not occurred (F / FMSY < 1), so it will recover in due course if the fishing intensity is continued at the existing level.
According to Figure
Simulated biomass time series of the fourteen specified high-commercial demersal fish species with 95% confidence intervals are considerable as shown in Figure
According to the above-described results of the Kobe diagrams, in which all stocks are in critical condition and are threatened with extinction, the trend plots of simulated biomass confirm the previous results, due to the Biomass not having a good increasing trend. As the charts in Figure
Conclusion
In summary, the Bayesian state-space GSPM under the MCMC algorithm was used to assess the stocks and provide fisheries management reference points for the fourteen high-commercial demersal fish species in the coastal domain of the study area. The authority of simulations about the models’ parameters and fisheries management reference points were approved by common diagnostic convergence tests. All the assessed fish stocks encountered were overfished and being overfished. The assessment outcomes, which reveals that the stock statuses of all targeted fish species were deteriorating indicates that the available fishery management strategies of small-scale fishery in the study area were not enough and new strategies associated with sustainable management were necessary.
As mentioned in the introduction, the scientific precise stock assessments have not been undertaken in Iran, especially on the southern coastal areas (such as the current study area) because of insufficient and limited information. Accordingly, it is one of the important reasons for the inefficiency of the available fishery management and conservation strategies for sustaining the studied fish species population status in the current study area. This reason is due to lack of applicable information for fisheries management and conservation planning (such as fisheries management reference points, biomass, harvest rate and stock status) that can be obtained from scientific precise stock assessments. Thus, in the short-term, the transferring of the obtained stock assessment outcomes of the current study to fisheries managers, planners and all other activists (such as fishermen) can improve the available fisheries strategies and harvesting treatments to rebuild and improve the current bad situation of studied fish species population status. Additionally, in the long run, the recommended use of the obtained stock assessment outcomes (e.g. management and biological reference points, biomass, harvest rate, stock status) for future research in line with appropriate ecosystem-based fishery management will determine the best strategies for preventing overfishing, improving, sustaining and conserving the above-overfished stocks. Hence, the obtained stock assessment outcomes in a viability theory framework to investigate various fishing scenarios for the implementation of the sustainable fishery management in the small-scale fishery sector of the current study area were used providing the details of the recommended viability theory modelling as an appropriate ecosystem-based fishery management approach in our further works.
Acknowledgments
The authors are grateful to Dr. Peter Sheldon Rankin from The University of Queensland, Australia, for his useful and constructive comments on estimating the above-mentioned models. The authors thank the National Fishery Data Collection Unit of Iran Fisheries Organization, particularly Mr. Sabah Khorshidi Nergi, the head of National Fishery Data Collection Unit for providing the fishery data. The authors also acknowledge the Provincial Fisheries Department of Sistan and Baluchestan in Chabahar for providing complementary and description information about artisanal fishing vessels operating in the multispecies fishery of the study area and Offshore Fisheries Research Centre of Chabahar for its contribution in specifying fish species. Finally, the authors express their thanks to the anonymous reviewers for their comments that improved our manuscript.
References
- Ando T (2010) Bayesian model selection and statistical modeling. CRC Press.
- Arendt JD (1997) Adaptive intrinsic growth rates: An integration across taxa. The Quarterly Review of Biology 72(2): 149–177. https://doi.org/10.1086/419764
- Berkes F (2001) Managing small-scale fisheries: alternative directions and methods. IDRC.
- Berliner LM (1996) Hierarchical Bayesian time series models. Maximum entropy and Bayesian methods. Springer, 15-22.
- Bishop J (2006) Standardizing fishery-dependent catch and effort data in complex fisheries with technology change. Reviews in Fish Biology and Fisheries 16(1): 21–38. https://doi.org/10.1007/s11160-006-0004-9
- Brodziak J, Ishimura G (2011) Development of Bayesian production models for assessing the North Pacific swordfish population. Fisheries Science 77(1): 23–34. https://doi.org/10.1007/s12562-010-0300-0
- Buckland S, Newman K, Thomas L, Koesters N (2004) State-space models for the dynamics of wild animal populations. Ecological Modelling 171(1-2): 157–175. https://doi.org/10.1016/j.ecolmodel.2003.08.002
- Chaloupka M, Balazs G (2007) Using Bayesian state-space modelling to assess the recovery and harvest potential of the Hawaiian green sea turtle stock. Ecological Modelling 205(1-2): 93–109. https://doi.org/10.1016/j.ecolmodel.2007.02.010
- Chen Y, Chen L, Stergiou KI (2003) Impacts of data quantity on fisheries stock assessment. Aquatic Sciences 65(1): 92–98. https://doi.org/10.1007/s000270300008
- Cheung WW, Sumaila UR (2015) Economic incentives and overfishing: A bioeconomic vulnerability index. Marine Ecology Progress Series 530: 223–232. https://doi.org/10.3354/meps11135
- Clark JS, Gelfand AE (2006) Hierarchical modelling for the environmental sciences: statistical methods and applications. Oxford University Press on Demand.
- Costello C, Ovando D, Hilborn R, Gaines SD, Deschenes O, Lester SE (2012) Status and solutions for the world’s unassessed fisheries. Science 338(6106): 517–520. https://doi.org/10.1126/science.1223389
- De Valpine P, Hastings A (2002) Fitting population models incorporating process noise and observation error. Ecological Monographs 72(1): 57–76. https://doi.org/10.1890/0012-9615(2002)072[0057:FPMIPN]2.0.CO;2
- Fletcher R (1978) On the restructuring of the Pella-Tomlinson system. Fish Bulletin 76: 515–521.
- Gelman A, Carlin JB, Stern HS, Rubin DB (2014) Bayesian data analysis. Chapman & Hall/CRC Boca Raton, FL, USA.
- Gelman A, Meng X-L (2004) Applied Bayesian modeling and causal inference from incomplete-data perspectives. John Wiley & Sons.
- Geweke J (1991) Evaluating the accuracy of sampling-based approaches to the calculation of posterior moments. Federal Reserve Bank of Minneapolis, Research Department Minneapolis, MN, USA.
- Gilks WR, Richardson S, Spiegelhalter DJ (1996) Introducing markov chain monte carlo. Markov chain Monte Carlo in practice 1: 19.
- Haddon M (2010) Modelling and quantitative methods in fisheries. CRC press.
- Harley SJ, Myers RA, Dunn A (2001) Is catch-per-unit-effort proportional to abundance? Canadian Journal of Fisheries and Aquatic Sciences 58(9): 1760–1772. https://doi.org/10.1139/f01-112
- Heidelberger P, Welch PD (1983) Simulation run length control in the presence of an initial transient. Operations Research 31(6): 1109–1144. https://doi.org/10.1287/opre.31.6.1109
- Hilborn R, Walters CJ (1992) Quantitative fisheries stock assessment: Choice, dynamics and uncertainty. Reviews in Fish Biology and Fisheries 2(2): 177–178. https://doi.org/10.1007/BF00042883
- Jiao Y, Cortés E, Andrews K, Guo F (2011) Poor‐data and data‐poor species stock assessment using a Bayesian hierarchical approach. Ecological Applications 21(7): 2691–2708. https://doi.org/10.1890/10-0526.1
- Jiao Y, Neves R, Jones J (2008) Models and model selection uncertainty in estimating growth rates of endangered freshwater mussel populations. Canadian Journal of Fisheries and Aquatic Sciences 65(11): 2389–2398. https://doi.org/10.1139/F08-141
- Kéry M, Schaub M (2011) Bayesian population analysis using WinBUGS: a hierarchical perspective. Academic Press.
- Kinas PG (1996) Bayesian fishery stock assessment and decision making using adaptive importance sampling. Canadian Journal of Fisheries and Aquatic Sciences 53(2): 414–423. https://doi.org/10.1139/f95-189
- King R, Morgan B, Gimenez O, Brooks S (2009) Bayesian analysis for population ecology. CRC Press.
- Kuparinen A, Mäntyniemi S, Hutchings JA, Kuikka S (2012) Increasing biological realism of fisheries stock assessment: Towards hierarchical Bayesian methods. Environmental Reviews 20(2): 135–151. https://doi.org/10.1139/a2012-006
- Laloë F (1995) Should surplus production models be fishery description tools rather than biological models? Aquatic Living Resources 8(1): 1–16. https://doi.org/10.1051/alr:1995001
- Mäntyniemi SHP, Whitlock RE, Perälä TA, Blomstedt PA, Vanhatalo JP, Rincón MM, Kuparinen AK, Pulkkinen HP, Kuikka OS (2015) General state-space population dynamics model for Bayesian stock assessment. ICES Journal of Marine Science: Journal du Conseil 72(8): 2209–2222. https://doi.org/10.1093/icesjms/fsv117
- Matthew S (2003) Small-scale fisheries perspectives on an ecosystem-based approach to fisheries management. Responsible Fisheries in the Marine Ecosystem, FAO, Rome: 47–64.
- McAllister MK, Kirkwood G (1998) Using Bayesian decision analysis to help achieve a precautionary approach for managing developing fisheries. Canadian Journal of Fisheries and Aquatic Sciences 55(12): 2642–2661. https://doi.org/10.1139/f98-121
- Meyer R, Millar RB (1999) BUGS in Bayesian stock assessments. Canadian Journal of Fisheries and Aquatic Sciences 56(6): 1078–1087. https://doi.org/10.1139/f99-043
- Millar RB (2002) Reference priors for Bayesian fisheries models. Canadian Journal of Fisheries and Aquatic Sciences 59(9): 1492–1502. https://doi.org/10.1139/f02-108
- Montenegro C, Branco M (2016) Bayesian state-space approach to biomass dynamic models with skewed and heavy-tailed error distributions. Fisheries Research 181: 48–62. https://doi.org/10.1016/j.fishres.2016.03.021
- Neiland AE, Béné C (2013) Poverty and small-scale fisheries in West Africa. Springer Science & Business Media.
- Osio GC, Orio A, Millar CP (2015) Assessing the vulnerability of Mediterranean demersal stocks and predicting exploitation status of un-assessed stocks. Fisheries Research 171: 110–121. https://doi.org/10.1016/j.fishres.2015.02.005
- Pakzad HR, Pasandi M, Soleimani M, Kamali M (2014) Distribution and origin of heavy metals in the sand sediments in a sector of the Oman Sea (the Sistan and Baluchestan province, Iran). Quaternary International 345: 138–147. https://doi.org/10.1016/j.quaint.2014.03.031
- Parent E, Rivot E (2012) Introduction to hierarchical Bayesian modeling for ecological data. CRC Press.
- Pauly D (2006) Major trends in small-scale marine fisheries, with emphasis on developing countries, and some implications for the social sciences. Maritime Studies 4: 7–22.
- Pella JJ, Tomlinson PK (1969) A generalized stock production model. Inter-American Tropical Tuna Commission.
- Plummer M, Best N, Cowles K, Vines K (2006) CODA: Convergence diagnosis and output analysis for MCMC. R News 6: 7–11.
- Punt AE, Hilborn RAY (1997) Fisheries stock assessment and decision analysis: The Bayesian approach. Reviews in Fish Biology and Fisheries 7(1): 35–63. https://doi.org/10.1023/A:1018419207494
- Punt AE, Smith DC, Smith ADM (2011) Among-stock comparisons for improving stock assessments of data-poor stocks: The “Robin Hood” approach. ICES Journal of Marine Science: Journal du Conseil 68(5): 972–981. https://doi.org/10.1093/icesjms/fsr039
- Punt AE, Szuwalski C (2012) How well can F MSY and B MSY be estimated using empirical measures of surplus production? Fisheries Research 134: 113–124. https://doi.org/10.1016/j.fishres.2012.08.014
- Quinn TJ, Deriso RB (1999) Quantitative fish dynamics. Oxford University Press.
- Rankin PS, Lemos RT (2015) An alternative surplus production model. Ecological Modelling 313: 109–126. https://doi.org/10.1016/j.ecolmodel.2015.06.024
- Salas S, Chuenpagdee R, Seijo JC, Charles A (2007) Challenges in the assessment and management of small-scale fisheries in Latin America and the Caribbean. Fisheries Research 87(1): 5–16. https://doi.org/10.1016/j.fishres.2007.06.015
- Schaefer MB (1957) A study of the dynamics of the fishery for yellowfin tuna in the eastern tropical Pacific Ocean. Inter-American Tropical Tuna Commission.
- Seaman JW III, Seaman Jr JW, Stamey JD (2012) Hidden Dangers of Specifying Noninformative Priors. The American Statistician 66(2): 77–84. https://doi.org/10.1080/00031305.2012.695938
- Spiegelhalter DJ, Best NG, Carlin BP, Van Der Linde A (2002) Bayesian measures of model complexity and fit. Journal of the Royal Statistical Society. Series B, Statistical Methodology 64(4): 583–639. https://doi.org/10.1111/1467-9868.00353
- Sturtz S, Ligges U, Gelman A (2010) R2OpenBUGS: a package for running OpenBUGS from R. http://wwwcranrprojectorg/web/packages/R2OpenBUGS/vignettes/R2OpenBUGSpdf
- Thomas A, O’Hara B, Ligges U, Sturtz S (2006) Making BUGS open. R News 6: 12–17.
- Valinassab T, Adjeer M, Momeni M (2010) Biomass estimation of demersal fishes in the Persian Gulf and Oman Sea by swept area method. Final Report. Iranian Fisheries Research Organization Press, Tehran.
- Valinassab T, Daryanabard R, Dehghani R (2003) Monitoring of demersal resources by swept area method in the Oman Sea waters. Final Report. Tehran, Iran
- Valinassab T, Daryanabard R, Dehghani R, Pierce GJ (2006) Abundance of demersal fish resources in the Persian Gulf and Oman Sea. Journal of the Marine Biological Association of the United Kingdom 86(06): 1455–1462. https://doi.org/10.1017/S0025315406014512
- Valinassab T, Dehghani R, Khorshidian K (2005) Biomass estimation of demersal resources in the Persian Gulf and Gulf of Oman by swept area method. Final Report Iranian Fisheries Research Organization (IFRO), Tehran, 121 pp.
- Wikle CK, Berliner LM, Cressie N (1998) Hierarchical Bayesian space-time models. Environmental and Ecological Statistics 5(2): 117–154. https://doi.org/10.1023/A:1009662704779
- Xiao Y (1998) Two simple approaches to use of production models in fish stock assessment. Fisheries Research 34(1): 77–86. https://doi.org/10.1016/S0165-7836(97)00064-7
- Ye Y, Dennis D (2009) How reliable are the abundance indices derived from commercial catch-effort standardization? Canadian Journal of Fisheries and Aquatic Sciences 66(7): 1169–1178. https://doi.org/10.1139/F09-070
- Yu H, Jiao Y, Carstensen LW (2013) Performance comparison between spatial interpolation and GLM/GAM in estimating relative abundance indices through a simulation study. Fisheries Research 147: 186–195. https://doi.org/10.1016/j.fishres.2013.06.002
- Zhu X, Day AC, Carmichael TJ, Tallman RF (2014) Hierarchical Bayesian Modeling for Cambridge Bay Arctic Char, Salvelinus Alpinus (L.), Incorporated with Precautionary Reference Points.
Appendix 1
Procedure of prior distributions functions used for parameters of all Bayesian state-space GSPM
Log-normal distribution procedure for intrinsic growth rate (r)
Standard Deviation = x
Average of Intrinsic Growth Rate (r) = y
Precision of Prior = 1/log(1 + x^2)
Average of Prior = log(y) – (0.5/ Precision of Prior)
r ~ dlnorm (Average of prior, Precision of prior)
Inverse Gamma Distribution Procedure for the Process and Observation Noise Variances
Shape Parameter = x
Scale Parameter=y
Gamma ~ dgamma (x, y)
Inverse-Gamma = 1/Gamma
Log-Normal Distribution Procedure for Initial Relative Biomass P[1]
B0=the Biomass in First Time
Kmin=minimum carrying capacity is considered equal to minimum Historical catches.
Kmax= maximum carrying capacity is considered equal to ten times the minimum Historical catches.
Pi ~ dunif(B0/Kmax, B0/Kmin)
isigma= Inverse Gamma Distribution for process noise variances
Pm[1] <- log(Pi)
P[1] ~ dlnorm(Pm[1], isigma)
Appendix 2
Trace plots for Bayesian state-space GSPM parameters (r, K, q, z, (sigma), and (tau)) for fourteen studied demersal fish species. Consider figures A1–14.













