Research Article |
|
Corresponding author: Bruno Petriccione ( b.petriccione@gmail.com ) Academic editor: Michele Freppaz
© 2019 Bruno Petriccione, Alessandro Bricca.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Petriccione B, Bricca A (2019) Thirty years of ecological research at the Gran Sasso d’Italia LTER site: climate change in action. In: Mazzocchi MG, Capotondi L, Freppaz M, Lugliè A, Campanaro A (Eds) Italian Long-Term Ecological Research for understanding ecosystem diversity and functioning. Case studies from aquatic, terrestrial and transitional domains. Nature Conservation 34: 9-39. https://doi.org/10.3897/natureconservation.34.30218
|
Abstract
Since 1986, vegetation monitoring of alpine plant communities has been performed at the Gran Sasso d’Italia LTER site (https://deims.org/c0738b00-854c-418f-8d4f-69b03486e9fd) in the Central Apennines, through phytosociological relevés and abundance and coverage estimation of the vascular flora at fine scale. The monitoring activities for abiotic parameters regard air and soil temperatures, rainfall, snowfall and snow cover persistence.
A comparative analysis of changes in species composition, life forms, life strategies and morpho-functional types allowed recognition of dynamical processes (fluctuation and degeneration) and an increase in stress- and drought-tolerant and ruderal species, probably linked to a general process of climate change.
A trend of variation forced by increasing drought was recorded in high-mountain plant communities, normally within a dynamic fluctuation process. There has been a 50–80% change in species composition with respect to the total number of species observed over the years. Whereas the total number of species has increased in all communities, in high-mountain mesic grassland 20% of sensitive species have completely disappeared. Early signs of a degeneration process were already discernible after seven years: such signs are more evident in snow-dependent communities, with a quantitative increase in more thermophilic and drought-tolerant species and a parallel decrease in more mesic, cryophilic and competitive species. In particular, the following phenomena have been recorded in high-mountain mesic grassland, in agreement with predicted or observed phenomena in other Alpine or Arctic areas: (a) coverage increase (or appearance) of ruderal and stress- and drought-tolerant species; (b) coverage decrease (or disappearance) of cryophilic, mesic and competitive species.
These short-term changes could lead, in the medium- or long-term, to a disgregation process affecting the high elevation plant communities of the Apennines (including the local extinction of most of the cold-adapted species), due to their very low resilience. The phenomena described may be linked to the observed climate change which occurred during the last century (in particular in the last 50 years) in the Apennines, consisting mainly, in the mountains, of a strong reduction in the duration of snow-cover and an increase in mean and minimum annual temperatures.
Keywords
LTER, climate change, alpine plant communities, Apennines, Braun-Blanquet approach
Introduction
According to the latest
Observed global climate warming in the 20th century (+0.7 °C in the last 60 years, +0.1 °C per decade in the same period,
As far as the Central Apennines (Italy) is concerned, data collected at some key meteorological stations show a considerable recent decrease in precipitation (especially in spring and autumn) and a small increase in average temperature (
This paper is based on continuous ecological observations performed at an alpine research site (“Gran Sasso d’Italia”, https://deims.org/c0738b00-854c-418f-8d4f-69b03486e9fd) established in 1986 and joining the LTER Italy network in 2007. The longest data series of vegetation data in the Apennines is available at this site, including up to 33 years of observations at community level. At European level, only data at species level are available for so long a time, except for an area in the Italian Alps (Stelvio National Park) where a 50-year data series is available (
Materials and methods
Study area
The LTER site EU IT 01-003-T “Appennino Centrale: Gran Sasso d’Italia”, established in 1985, covers an area of ca. 0.5 km2, with an elevation range from 2130 to 2385 m a.s.l. (approx. latitude 42°26N and longitude 13°33E, Suppl. material
The climate is Mediterranean-mountain (
Since 1986, the same ecologists (first from the Universities of Rome and L'Aquila, from 1999 the Corpo Forestale dello Stato (National Forest Service) and since 2017 the Biodiversity and Park Protection Department of the Carabinieri) have been continuously studying the state of the high elevation vegetation, analysing all the plant species present in permanent plots where surveys are carried out once a year. Since 2013, the microclimate has also been studied directly using automatic temperature measurement devices installed in the soil.
The site consists of high elevation plant communities above the timberline, in the central Mediterranean basin, along the Apennine mountain range, included in the alpine and Mediterranean high-mountain altitudinal belts (
Climate data
Climate data were collected at the official Campo Imperatore meteorological station (2135 m a.s.l.), located in the Gran Sasso d’Italia mountains, with over 70 years of standard precipitation and temperature observations (since 1942). Data on snow precipitation and the height of snow cover in winter were collected at the non-automatic METEOMONT station (managed by the Carabinieri, formerly the National Forest Service) at the same location, from 1988 to 2016. Total winter snow precipitation has been calculated on the basis of METEOMONT data (daily visual observations of height of new snow cover, in cm, assuming 1 cm of snow equal to 1 mm of liquid precipitation).
Continuous soil microclimate observations have been made at the site since 2013, by means of two data loggers in the soil (-10 cm depth), one for each community, according to the GLORIA methodology (
Analysis at community level
For each of the six plots, 100 m2 in area, a phytosociological relevé was performed (according to the Braun-Blanquet approach) in 1993 and repeated 6, 8 and 15 years later, in 1999, 2001 and 2008, on the same fixed surface area of the entire plot. Since 2008, data collection has been yearly, including three plots per community. In one case (Pediculari-Seslerietum), a relevé had also been performed seven years earlier (in 1986), even if in only one plot. All data are reported in Suppl. material
The ecology and distribution range of each plant community are described according to
Species composition changes were calculated by simply comparing year by year, each year with the original species composition (at 1986 or 1993) and with all species occurring during the 25–33 years of observation.
A temporal trend analysis of species composition, life forms (
To analyse changes over time at community level, the species cover value for each plot (
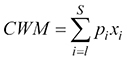
where xi is the mean trait values and pi is the relative abundance of the i-th species. To calculate the CWM with trait (hereafter CWMtrait) the matrixes “relevés x species (cover %)” and “species x trait values” were used. This index can be adequately used to quantify the relationship between a given trait and certain environmental conditions. The variation in life strategies was also approached using the above equation, multiplying the matrix “relevés x species (cover %)” with the matrix “species x Grime’s strategy” using the same equation of
Finally, an analysis of the change over time of the community’s ecological features was performed using the ecological bioindicator values (hereafter BV) for the Italian flora (
Analysis at species level
At species level, changes in the cover of dominant and other selected species were analysed for both communities along a continuous temporal gradient (as performed by
The same statistical method as described above was employed and a non-parametric linear regression model was used when the assumptions for the parametric model were not satisfied.
All the analyses were computed using R software (version 3.4.4, R Foundation for Statistical Computing,
Results
Climate
Analysis of precipitation and temperature in the period 1950–2014 (65 years) shows (Fig.

Changes in climate features during the period 1950–2014 at the Gran Sasso d’Italia LTER site (a mean annual air temperature (MAT) b mean annual total precipitation (SAP) c mean winter snow depth (MSC) for the period 1988–2016 d total winter snow precipitation (MSP) for the period 1988-2016 (SWE, mm); *: regression performed using non-parametric linear regression analysis; ns: non-significant).
For snow data, a significant variation (decrease) was found for snow precipitation only (Fig.
The continuous soil microclimate observations performed at the site (Figs

Daily Seasonal and annual changes in soil temperature (°C, recorded on a hourly bases -10 cm depth), at the dry grassland plots (Pediculari-Seslerietum) in the period May 2013-July 2018.

Community level
Species composition changed by 10–15% per year in the period 1993–2017 (25 years) in both communities; the rate of change is also quite similar if the observation period is extended to a total of 32 years (possible only in the case of the Pediculari-Seslerietum). Long-term analysis, however, shows different trends: the dry grasslands (Pediculari-Seslerietum) have not lost any species, but have gained 53% of the total number of species occurring during the 32 years of observation; the mesic grasslands (Luzulo-Festucetum), on the other hand, have lost 20% of the original species (at 1993) and gained a very high number (81%) of the total number of species occurring during the 25 years of observation. Despite these very relevant changes in species composition, both communities maintain the same total plant coverage values over time (60–70% for Pediculari-Seslerietum and 90–95% for Luzulo-Festucetum).
The results show different biodiversity features of the two communities (taxonomic and functional) changing over time (Figs

Changes in species diversity traits during the period 1986–2017 (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in species diversity traits during the period 1993–2017 (Luzulo-Festucetum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).
As regards morpho-functional features (Figs

Changes in morpho-functional traits during the period 1986–2017 (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1986–2017 (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1986–2017 (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1986–2017 (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1993–2017 (Luzulo-Festucetum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1993–2017 (Luzulo-Festucetum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1993–2017 (Luzulo-Festucetum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in morpho-functional traits during the period 1993–2017 (Luzulo-Festucetum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).
As far as the analysis of Grime’s life strategies (Figs

Changes in life strategies during the period 1986–2017 (Pediculari-Seslerietum plots) (S: stress-tolerant; R: ruderal; C: competitive) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in life strategies during the period 1993)-2017 (Luzulo-Festucetum plots) (S: stress-tolerant; R: ruderal; C: competitive) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).
As regards ecological indicators, generating ecograms of the two plant communities (Suppl. material

Changes in ecological bioindicator values during the period 1986–2017 (Pediculari-Seslerietum plots) (a): light availability; b): thermophile behaviour; c): continentality gradient; d): pH gradient; e): nitrogen availability; f): soil moisture) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Changes in ecological bioindicator values during the period 1993–2017 at the Gran Sasso d’Italia LTER site (Luzulo-Festucetum plots) (a): light availability; b): thermophile behaviour; c): continentality gradient; d): pH gradient; e): nitrogen availability; f): soil moisture) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).
Species level
The results of the analysis at species level highlighted a preferential response for the species belonging to the Luzulo-Festucetum, with a lack of response for the selected species of the Pediculari-Seslerietum. In detail, a significant decreasing trend has been found for the former dominant species Festuca violacea and a significative increasing trend for the new dominant species Trifolium pratense and the invaders Carex caryophyllea (even if not significant) which have only colonised the community since 2008 (Figs

Changes in the coverage values of some selected species (Pediculari-Seslerietum plots) (*: regression performed using non-parametric linear regression analysis; ns: non-significant).

Discussion
Climate data are concordant with those presented in
Decreased snowfall and snow persistence (and increased variability), together with the variability of total precipitation, increase the ecological stress at the research site, as confirmed by interpretation of the soil microclimate data collected during the last five years: the recorded frequent frost episodes, connected with the absence of protective snow cover, will expose the biocenoses to very dangerous frost stress. The official meteorological station at the same location (with sensors well above the soil surface) confirms these data, with 167 frost days in winter 2013–2014: only 6 days more than data related to the Pediculari-Seslerietum (which therefore remains exposed to frost for a few days only), but 10 days less than data for the Luzulo-Festucetum (which conserves the protective snow cover for longer). Decreased snowfall is in agreement with previous observations in the Apennines (
The low yearly rate of species turnover observed at the site (confirming the preliminary results from
Although both communities preserve the same values of total plant coverage over time, in the case of the Luzulo-Festucetum the former dominant species (Festuca violacea) lost its past role, due to the invasion and expansion of an opportunistic species (Trifolium pratense). These results are in good agreement with those found in the Central Italian Alps in the subalpine, alpine and nival belts over the last 50 years (
The significant changes of morpho-functional traits, noted in both communities, are interpretable as the effects of a decrease of snow cover and an increase in temperature and drought stress (
Regarding the Luzulo-Festucetum mesic grassland, on the other hand, we found a more complex response to the variation over time. In line with the trend of increasing drought, we noted a significant decrease in species without hairs and a significant increase in species with sparse hairs. The observed increase of leaf width is apparently in contrast with increasing drought stress. However, this pattern may be the result of an increase in compound leaves, given that for these species the leaf, and not the leaflet, was measured. An increase in compound leaves along the aridity gradient has been noted (
The significant changes in life strategies, with an increase in species with a stress-tolerant strategy for the mesic community, are characteristic of biocenoses with increased ecological stress, undergoing changes towards adaptation to an increased drought. These results are in agreement with observations made in the Alps (
The progressive narrowing of the Luzulo-Festucetum ecogram over time confirms the correlation between the observed warmth and drought trends and the related changes in the sensitive mesic plant community. Our results exclude a possible effect on changes in vegetation caused by modified land use or nitrogen accumulation in the soil (as shown by the related ecogram, with a clear decrease in nitrogen availability indicators over time), unlike the assumptions of
Dry grassland shows higher resistance to an increased drought than mesic grassland. To improve the reliability of the findings, a regression model (non-parametric where assumptions were not met) removing data collected before 2008 (with no replicates) was also performed (Suppl. material
Conclusions
The warming trend at global level is confirmed and reinforced by data related to the LTER site “Gran Sasso d’Italia”: the mean annual temperature has increased by 1.7 °C over the last 65 years, corresponding to an average increase per decade of +0.26 °C. This is more than double the same values at global level (+0.7 °C in the last 60 years and +0.1 °C per decade,
This exceptional warming in alpine areas, together with a decrease in total precipitation (as recognized for the Central Apennines as a whole, even if not significant at the site) and snowfall (significant at the site), an increase in climate inter-annual variability and extreme events, and a frequent lack of snow cover, are the combined drivers of the intense species turnover observed, occurring over the last 30 years in all the biocenoses studied, although more marked in snow-dependent communities. A quantitative increase in more thermophilic and stress- and drought-tolerant species and a parallel decrease in more mesic, cold adapted and competitive species have been clearly detected. These results confirm the preliminary assumptions provided in
Ecological indicators demonstrate that the key factor in the ecological changes of the alpine biocenoses studied is drought, associated with the combined action of temperature increase, precipitation decrease and lack of snow cover and precipitation.
The two communities studied react in different ways to these abiotic drivers: (1) the Pediculari-Seslerietum dry grassland, highly resistant and well adapted to drought, frost and drastic temperature ranges, shows very slow or no changes over time (in accordance with the results of
The fluctuation stage typical of these primary alpine plant communities seems to be changing toward a dynamical tendency of degeneration, with an important disgregation of the community due to deterioration of the ecological connections: as in the Central Alps, this process can lead to an ecological vacuum or a disequilibrium state in the biocenoses (
In conclusion, our results enable us to answer the four questions listed in the introduction: a) plant communities are significantly changing over time, more for mesic grassland and less for dry grassland; b) toward a disequilibrium state; c) species are responding in different ways, altering the intra-community ecological connections; d) there is a relationship between the changes in the features of the communities and the predicted and the observed changes in the temperature and precipitation regimes.
Additional long-term observations over the next decades are, in any case, required to confirm the hypothesis of a cause-effect relationship between climate change and changes in plant communities and to exclude natural and unknown fluctuations.
The combined monitoring of vegetation (composition and structure) and temperature at high elevation will provide updated data on the processes currently underway on the high summits of the Apennines and will guide the local in-situ policies to conserve the associated plant communities and threatened species.
Acknowledgments
B.P. thanks Sarah Gregg, Monia Marrone, Linda Brucculeri, Claudia Cindolo and Cristiana Cocciufa for their sharing and valued contribution in the field work. Further thanks to Sarah Gregg for the mother tongue revision. Research performed also within the Virtual Access activities of the project eLTER-H2020 (GA654359) in collaboration with CNR-IRET.
References
- Baldoni M, Biondi E, Frattaroli AR (1999) Caratterizzazione bioclimatica del Gran Sasso d’Italia. Braun-Blanquetia 16: 7–20.
- Biondi E, Blasi C, Burrascano S, Casavecchia S, Copiz R, Del Vico E, Galdenzi D, Gigante D, Lasen C, Spampinato G, Venanzoni R, Zivkovic L (2009) Manuale Italiano di interpretazione degli habitat della Direttiva 92/43/CEE. Società Botanica Italiana. Ministero dell’Ambiente e della tutela del territorio e del mare. http://vnr.unipg.it/habitat
- Blasi C (1996) Un approccio fitoclimatico allo studio dei cambiamenti climatici in Italia. Atti – Società Italiana di Ecologia 17: 34–39.
- Braun-Blanquet J (1932) Plant Sociology. New York.
- Braun-Blanquet J (1955) Die Vegetation des Piz Languard, ein Maßstab für Klimaänderungen. Svensk Botanisk Tidskrift 49: 1–9.
- Braun-Blanquet J (1964) Pflanzensoziologie. Grundzuge der Vegetationskunde. Springer, Wien and New York. https://doi.org/10.1007/978-3-7091-8110-2
- Braun-Blanquet J (1957) Ein Jahrhundert Florenwandel am Piz Linard (3414 m). Extrait du Bull. Jard.Botan. Bruxelles, Vol. Jubil. W. Robyns, Comm. S.I.G.M.A. 137: 221–232.https://doi.org/10.2307/3666959
- Brucculeri R, Petriccione B (1994) Adattamenti e microclima delle comunità vegetali d'alta quota degli Appennini (Italia). Revue Valdôtaine d'Histoire Naturelle 48(suppl.): 131–141.
- Brunetti M, Buffoni L, Maugeri M, Nanni T (2000a) Trend of minimum and maximum daily temperature in Italy from 1865 to 1996. Theoretical and Applied Climatology 66: 49–60. https://doi.org/10.1007/s007040070032
- Brunetti M, Maugeri M, Nanni T (2000b) Variations of temperature and precipitation in Italy from 1866 to 1995. Theoretical and Applied Climatology 65: 165–174. https://doi.org/10.1007/s007040070041
- Bucchignani E, Montesarchio M, Zollo AL, Mercogliano P (2016) High‐resolution climate simulations with COSMO‐CLM over Italy: performance evaluation and climate projections for the 21st century. International Journal of Climatology 36: 735–756. https://doi.org/10.1002/joc.4379.
- Buffoni L, Maugeri M, Nanni T (1999) Precipitation in Italy from 1833 to 1996. Theoretical and Applied Climatology 63: 33–40.https://doi.org/10.1007/s007040050089
- Camenisch M (2002) Veränderungen der Gipfelflora im Bereich des Schweizerischen Nationalparks: Ein Vergleich über die letzen 80 Jahre. Jahresber. Nat.forsch. Ges. Graubünden 111: 27–37.
- Cannone N, Pignatti S (2014) Ecological responses of plant species and communities to climate warming: upward shift or range filling processes? Clim. Change 123: 201–214. https://doi.org/10.1007/s10584-014-1065-8
- Cannone N, Sgorbati S, Guglielmin M (2007) Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 5 (7): 360–365. https://doi.org/10.1890/1540-9295(2007)5[360:UIOCCO]2.0.CO;2
- Chapin FS III, Bret-Harte MS, Hobbie SE, Zhong H (1996) Plant functional types as predictors of transient responses of arctic vegetation to global change. J. Veg. Sci. 7: 347–358. https://doi.org/10.2307/3236278
- Clementi A (1995) L’evoluzione storica del territorio. In: Rolli GL, Romano B (Eds) Progetto Parco. Tutela e valorizzazione dell’ambiente nel comprensorio del Gran Sasso d’Italia. Andromeda Editrice, Teramo, 176–196.
- Dirnböck T, Dullinger S, Grabherr G (2003) A regional impact assessment of climate and land-use change on alpine vegetation. Journal of Biogeography 30: 401–429. https://doi.org/10.1046/j.1365-2699.2003.00839.x
- Edwards AC, Scalenghe R, Freppaz M (2007) Changes in the seasonal snow cover of alpine regions and its effect on soil processes: A review. Quaternary International 162–163: 172–181. https://doi.org/10.1016/j.quaint.2006.10.027
- Erschbamer B, Kiebacher T, Mallaun M, Unterluggauer P (2009) Short-term signals of climate change along an altitudinal gradient in the South Alps. Plant Ecol. 202: 79–89. https://doi.org/10.1007/s11258-008-9556-1
- Erschbamer B, Unterluggauer P, Winkler E, Mallaun M (2011) Changes in plant diversity revealed by long-term monitoring on mountain summits in the Dolomites (Italy). Preslia 83: 387–401. http://www.preslia.cz/P113Erschbamer.pdf
- Evangelista A, Frate L, Carranza ML, Attorre F, Pelino G, Stanisci A (2016) Changes in composition, ecology and structure of high-mountain vegetation: A re-visitation study over 42 years. AoBPlants 8: 1–11. https://doi.org/10.1093/aobpla/plw004
- Falcucci A, Maiorano M, Boitani L (2007) Changes in land-use/land-cover patterns in Italy and their implications for biodiversity conservation. Landscape Ecol 22: 617–631. https://doi.org/10.1007/s10980-006-9056-4
- Falinski B (1986) Vegetation dynamics in temperate lowland primeval forest. Ecological studies in Bialowieza forest. Geobotany 8: 1–537. https://doi.org/10.1007/978-94-009-4806-8_1
- Falinski B (1989) Le temp et l'éspace dans les recherches écologiques sur le dynamisme de la végétation. Giorn. Bot. Ital. 123: 81–107. https://doi.org/10.1080/11263508909430249
- Frate L, Carranza ML, Evangelista A, Stinca A, Schaminée JHJ, Stanisci A (2018) Climate and land use change impacts on Mediterranean high-mountain vegetation in the Apennines since the 1950s. Plant Ecology & Diversity, 11: 85–96. https://doi.org/10.1080/17550874.2018.1473521
- Furrer E, Furnari F (1960) Ricerche introduttive sulla vegetazione di altitudine del Gran Sasso d’Italia. Boll. Ist. Bot. Univ. Catania 2-2: 143–201.
- Garnier E, Cortez J, Billès G, Navas ML, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Tousaint JP (2004) Plant functional ecology markers capture ecosystems properties during secondary succession. Ecology 85: 2630–2637. https://doi.org/10.1890/03-0799
- Givnish TJ (1978) Adaptive significance of compound leaves with particular reference to tropical trees. In: Tomlinson PB, Zimmerman MH (eds.). Tropical Trees as Lil'ing Sy.l/em~. Cambridge University Press, Cambridge.
- Givnish TJ (1987) Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints. New phytologist 106: 131–160. https://doi.org/10.1111/j.1469-8137.1987.tb04687.x
- Gottfried M, Pauli H, Futschik A, Akhalkatsi M, Barančok P, Benito Alonso JL, Coldea G, Dick J, Erschbamer B, Fernández Calzado MR, Kazakis G, Krajči J, Larsson P, Mallaun M, Michelsen O, Moiseev D, Moiseev P, Molau U, Merzouki A, Nagy L, Nakhutsrishvili G, Pedersen B, Pelino G, Puscas M, Rossi G, Stanisci A, Theurillat J-P, Tomaselli M, Villar L, Vittoz P, Vogiatzakis I, Grabherr G (2012) Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2: 111–115. https://doi.org/10.1038/nclimate1329
- Gottfried M, Pauli H, Reiter K (1999) A fine-scaled predictive model for changes in species distribution patterns of high mountain plants induced by climate warming. Diversity and Distributions 5: 241–251. https://doi.org/10.1046/j.1472-4642.1999.00058.x
- Gottfried M, Pauli H, Reiter K (2002) Potential effects of climate change on alpine and nival plants in the Alps. In: Körner C, Spehn EM (eds.) Mountain biodiversity, a global assessment. Parthenon Publishing, London, New York, 213–223.
- Grabherr G, Gottfried M, Gruber A (1995) Patterns and current changes in alpine plant diversity. In: Chapin III FS, Körner C (Eds) Arctic and alpine biodiversity: patterns, causes and ecosystem consequences, 113. Springer, Berlin, 167–181. https://doi.org/10.1007/978-3-642-78966-3_12
- Grabherr G, Gottfried M, Pauli H (1994) Climate effects on mountain plants. Nature, 369: 448–448. https://doi.org/10.1038/369448a0
- Grabherr G, Gottfried M, Pauli H (2001) Long-term monitoring of mountain peaks in the Alps. In: Burga CA, Kratochwil A (Eds) Biomonitoring: General and applied aspects on regional and global scales, 35. Tasks for Vegetation Science, Kluwer, Dordrecht, 153–177. https://doi.org/10.1007/978-94-015-9686-2_10
- Grime JP (1977) Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist 3: 1169–1194. https://doi.org/10.1086/283244
- Grime JP (1984) Towards a functional description of vegetation. In: White J, Beeftink J (Eds) Population Structure of Vegetation. Junk, The Hague. https://doi.org/10.1007/978-94-009-5500-4_21
- Grime JP (2006) Plant strategies, vegetation processes, and ecosystem properties. John Wiley & Sons, Toronto.
- Guisan A, Theurillat J-P (2000) Assessing alpine plant vulnerability to climate change: a modelling perspective. Integrated Assessment 1: 307–320. https://doi.org/10.1023/A:1018912114948
- Hofer HR (1992) Veränderungen in der Vegetation von 14 Gipfeln des Berninagebietes zwischen 1905 und 1985. Ber Geobot Inst ETH. Stift Rubel 58: 39–54.
- Holzinger B, Hülber K, Camenisch M (2008) Changes in plant species richness over the last century in the eastern Swiss Alps: elevational gradient, bedrock effects and migration rates. Plant Ecology 195: 179–196. https://doi.org/10.1007/s11258-007-9314-9
- Intergovernmental Panel on Climate Change (2001) Climate Change 2001: Impacts, Adaptation and Vulnerability. Cambridge Univ. Press, UK. https://www.ipcc.ch/ipccreports/tar/wg1/pdf/WGI_TAR_full_report.pdf
- Intergovernmental Panel on Climate Change (2007) Climate Change 2007: Impacts, Adaptation and Vulnerability. Cambridge Univ. Press, UK. https://www.ipcc.ch/pdf/assessment-report/ar4/wg2/ar4_wg2_full_report.pdf
- Intergovernmental Panel on Climate Change (2014) Climate Change 2014: Impacts, Adaptation and Vulnerability. Cambridge Univ. Press, UK. https://doi.org/10.1017/CBO9781107415416
- Klanderud K, Birks HJB (2003) Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. The Holocene 13: 1–6. https://doi.org/10.1191/0959683603hl589ft
- Knight CA, Ackerly DD (2003) Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: congeneric species from desert and coastal environments. New Phytologist 160(2): 337–347. https://doi.org/10.1046/j.1469-8137.2003.00880.x
- Körner C (1994) Impact of atmospheric changes on high mountain vegetation. In: Beniston M (Ed.) Mountain Environments in Changing Climates. Routledge Publ. Co., London.
- Körner C (1999) Alpine plant life. Functional plant ecology of high mountain ecosystems. Springer, Berlin. https://doi.org/10.1007/978-3-642-98018-3
- Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail, Functional Ecology 16: 545–556. https://doi.org/10.1046/j.1365-2435.2002.00664.x
- Lundquist JD, Lott F (2008) Using inexpensive temperature sensors to monitor the duration and heterogeneity of snow-covered areas. Water Resources Research 44. https://doi.org/10.1029/2008WR007035
- Mathez EA (2009) Climate Change. The Science of Global Warming and Our Energy Future. Columbia University Press. https://cup.columbia.edu/book/climate-change/9780231146425.
- Moiseev PA, Shiyatov SG (2003) Vegetation dynamics at the treeline ecotone in the Ural highlands, Russia. In: Nagy L, Grabher G, Körner C, Thompson DBA (Eds) Alpine biodiversity in Europe. A Europe-wide assessment of biological richness and change. 167. Springer, Berlin, 423–435. https://doi.org/10.1007/978-3-642-18967-8_26
- Parolo G, Rossi G (2008) Upward migration of vascular plants following a climate warming trend in the Alps. Basic and Applied Ecology 9: 100–107. https://doi.org/10.1016/j.baae.2007.01.005
- Pauli H, Gottfried M, Dirnböck T (2003a) Assessing the long-term dynamics of endemic plants at summit habitats. In: Nagy L, Grabherr G, Körner C, Thompson DB (Eds) Alpine Biodiversity in Europe – A Europe-wide assessment of biological richness and change. Springer, Berlin, 195–207. https://doi.org/10.1007/978-3-642-18967-8_9
- Pauli H, Gottfried M, Grabherr G (2001) High summits of the Alps in a changing climate. In: Walther G, Burga CA, Edwards PJ (Eds) “Fingerprints” of Climate Change: Adapted Behaviour and Shifting Species Ranges. Kluwer Academic/Plenum, New York, 139–149. https://doi.org/10.1007/978-1-4419-8692-4_9
- Pauli H, Gottfried M, Grabherr G (2003b) The Piz Linard (3411m), the Grisons, Switzerland – Europe's oldest mountain vegetation study site. In: Nagy L, Grabherr G, Körner C, Thompson DBA (Eds) Alpine Biodiversity in Europe – A Europe-wide assessment of biological richness and change. Springer, Berlin, 443–448. https://doi.org/10.1007/978-3-642-18967-8_28
- Pauli H, Gottfried M, Hohenwallner D (2004) The GLORIA field manual. Multi-Summit approach. European Commission, DG Research, EUR 21213, Office for Official Publications of the European Communities, European Commission, Luxembourg, 85 pp.
- Pauli H, Gottfried M, Reiter K (2007) Signals of range expansions and contractions of vascular plants in the high Alps: observations (1994–2004) at the GLORIA master site Schrankogel, Tyrol, Austria. Global Change Biology 13: 147–156. https://doi.org/10.1111/j.1365-2486.2006.01282.x
- Petriccione B (1994) Ecologia e fitogeografia delle praterie di altitudine a Sesleria tenuifolia degli Appennini (Italia). Rev. Valdotaine Hist. Nat. suppl. 48: 287–296.
- Petriccione B (1995) Land ecosystems sensitive to climate change (possible impact). In: Ministry of Environment. First Italian National Communication to the Framework Convention on Climate Change, 137–148.
- Petriccione B (2005) Short-term changes in key plant communities of Central Apennines (Italy). Acta Botanica Gallica 152: 545–561. https://doi.org/10.1080/12538078.2005.10515513
- Petriccione B (2012) Appennino Centrale: Gran Sasso d’Italia. In: Bertoni R (Ed.) La Rete Italiana per la Ricerca Ecologica a Lungo Termine (LTER-Italia): situazione e prospettive dopo un quinquennio di attività (2006–2011). ARACNE Editrice, Roma, 41–43. http://www.ise.cnr.it/crypta/ebooks/La%20rete%20italiana%20LTER.pdf
- Petriccione B, Carotenuto L, Crisanti L (1996) Ecosistemi terrestri particolarmente vulnerabili al cambiamento climatico: possibili impatti. Atti S.It.E. 17: 71–74.
- Petriccione B, Carotenuto L, Crisanti L (1998) Principali biocenosi terrestri vulnerabili al cambiamento climatico: stato attuale ed ipotesi sui cambiamenti a medio e lungo termine. Doc. Phytosoc. 28: 1097–1119.
- Petriccione B, Claroni N (1996) The dynamical tendencies in the vegetation of Velino massif (Abruzzo, Italy). Doc. Phytosoc. 16: 365–373.
- Petriccione B, Persia G (1995) Prodromo delle praterie di altitudine su calcare degli Appennini (classe Festuco-Seslerietea). Atti Conv. Lincei 115: 361–389.
- Pignatti S (1952) Fitogeografia. In: Cappelletti C, Trattato di Botanica. UTET, Torino.
- Pignatti S (1969) Arealtypen und die antstehung der apenninischen gebirgsflora. Mitt. Ostalp.-din. Pfalnzensoz. Arbeitsgem., Camerino.
- Pignatti S (2005) Valori di bioindicazione delle piante vascolari della flora d’Italia. Braun-Blanquetia 39: 3–97. http://www.scienzadellavegetazione.it/sisv/libreria/braun-blanquetia/BRBL39.pdf
- Pignatti S (2017–2019) Flora d’Italia. Edagricole-New Business Media, Bologna. 4 Vols.
- R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org
- Raunkiaer C (1934) Life forms of plants and statistical plant geography. Oxford.
- Rogora M, Frate L, Carranza ML, Freppaz M, Stanisci A, Bertani I, Bottarin R, Brambilla A, Canullo R, Carbognani M, Cerrato C, Chelli S, Cremonese I, Cutini M, Di Musciano M, Erschbamerm B, Godone D, Iocchi M, Isabellon M, Magnani A, Mazzola L, Morra di Cella U, Pauli H, Petey M, Petriccione B, Porro S, Psenner R, Rossetti G, Scotti A, Sommaruga R, Tappeiner U, Theurillat J-P, Tomaselli M, Viglietti D, Viterbi R, Vittoz P, Winkler M, Matteucci G (2018) Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Science of the Total Environment 624: 1429–1442. https://doi.org/10.1016/j.scitotenv.2017.12.155
- Romeo V, Scarpelli F (2001) Le condizioni meteonivometriche negli Appennini. Linea Ecologica 4: 44–53.
- Rossi G, Parolo G, Dellavedova R (2004) Gli organismi vegetali come bioindicatori dei cambiamenti climatici: il progetto GLORIA. Atti del convegno "Acque a Cremona" - Museo civico di Storia Naturale di Cremona, 81–94.
- Rossi G, Parolo G, Zonta LA, Crawford JA, Leonardi A (2006) Salix herbacea L. fragmented small population in the N-Apennines (Italy): response to human trampling disturbance. Biodiversity and Conservation 15: 3881–3893. hhttps://doi.org/10.1007/s10531-005-0607-9
- Schmid MO, Gubler S, Fiddes J, Gruber S (2012) Inferring snowpack ripening and melt-out from distributed measurements of near-surface ground temperatures. The Cryosphere 6: 1127–1139. https://doi.org/10.5194/tc-6-1127-2012
- Stanisci A, Frate L, Morra di Cella U, Pelino G, Petey M, Siniscalco C, Carranza ML (2016) Short-term signals of climate change in Italian summit vegetation: observations at two GLORIA sites. Plant Biosyst. 150: 227–235. https://doi.org/10.1080/11263504.2014.968232
- Stanisci A, Pelino G, Blasi C (2005) Vascular plant diversity and global change in central Apennine (Italy). Biodiversity and Conservation 14: 1301–1318. https://doi.org/10.1007/s10531-004-9674-6
- Stanisci A, Pelino G, Guisan A (2006) Cambiamenti climatici ed effetti sulla flora d'alta quota nel Parco Nazionale della Majella. In: Di Cecco M, Andrisano T (Eds) La biodiversità vegetale nelle aree protette in Abruzzo: studi ed esperienze a confronto. Parco Nazionale della Majella, Guardiagrele, 192–209.
- Steinbauer MJ, Jurasinski J-A G, Kulonen A, Lenoir J, Pauli H, Rixen C, Winkler M, Bardy-Durchhalter M, Barni E, Bjorkman AD, Breiner FT, Burg S, Czortek P, Dawes MA, Delimat A, Dullinger S, Erschbamer B, Felde VA, Fernández-Arberas O, Fossheim KF, Gómez-García D, Georges D, Grindrud ET, Haider S, Haugum SV, Henriksen H, Herreros MJ, Jaroszewicz B, Jaroszynska F, Kanka R, Kapfer J, Klanderud K, Kühn I, Lamprecht A, Matteodo M, Morra di Cella U, Normand S, Odland A, Olsen SL, Palacio S, Petey M, Piscová V, Sedlakova B, Steinbauer K, Stöckli V, Svenning J-C, Teppa G, Theurillat J-P, Vittoz P, Woodin SJ, Zimmermann NE, Wipf S (2018) Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 556: 231–234. https://doi.org/10.1038/s41586-018-0005-6
- Theurillat JP, Guisan A (2001) Potential impact of climate change on vegetation in the European Alps: a review. Climatic Change 50, 77-109. https://doi.org/10.1023/A:1010632015572
- Thuiller W, Lavorel S, Araujo MB (2005) Climate change threats to plant diversity in Europe. Proceedings of the National Academy of Sciences of the United States of America, 102: 8245–8250. https://doi.org/10.1073/pnas.0409902102
- Virtanen R, Eskelinen A, Gaare E (2003) Long-Term Changes in Alpine Plant Communities in Norway and Finland. In: Nagy L, Grabherr G, Körner C, Thompson DBA (Eds) Alpine biodiversity in Europe – A Europe-wide assessment of biological richness and change. Springer, Berlin, 411–422. https://doi.org/10.1007/978-3-642-18967-8_25
- Vittoz P, Bodin J, Ungricht S (2008) One century of vegetation change on Isla Persa, a nunatak in the Bernina massif in the Swiss Alps. J. Veg. Sci.19: 671–U628. https://doi.org/10.3170/2008-8-18434
- Wagner GJ, Wang E, Shepherd RW (2004) New approaches for studying and exploiting an old protuberance, the plant trichome. Annals of Botany 93(1): 3–11. https://doi.org/10.1093/aob/mch011
- Walther GR, Beißner S, Burga CA (2005) Trends in upward shift of alpine plants. J. Veg. Sci. 16: 541–548. https://doi.org/10.1111/j.1654-1103.2005.tb02394.x
- Welker JM, Bowman WD, Seastedt TR (2001) Environmental change and future directions in alpine research. In: Bowman WD, Seastedt R (Еds) Structure and function of an alpine ecosystem: Niwot Ridge, Colorado. Oxford Univ. Press, New York.
- Woodward FI, Cramer W (1996) Plant functional types and climatic changes: Introduction. J. Veg. Sci. 7: 306–308. https://doi.org/10.1111/j.1654-1103.1996.tb00489.x
- Zollo AL, Rillo V, Bucchignani E, Montesarchio M, Mercogliano P (2016) Extreme temperature and precipitation events over Italy: assessment of high‐resolution simulations with COSMO‐CLM and future scenarios. Int. J. Climatol. 36: 987–1004. https://doi.org/10.1002/joc.4401
Supplementary material
Supplementary tables and figures
Data type: measurements
Explanation note: Table S1. Pediculari-Seslerietum 1986–2017 (phytosociological releves). Table S2. Luzulo-Festucetum 1993–2017 (phytosociological releves). Figure S1. Geographical location of the LTER site “Gran Sasso d’Italia” (site area in pink; permanent plots indicated by red squares). Figure S2. Pedological profile of the Gran Sasso d’Italia Pediculari-Seslerietum community (soil depth in cm; from