Research Article |
|
Corresponding author: Douglas William Cirino ( douglaswcirino@hotmail.com ) Academic editor: Cristian-Remus Papp
© 2022 Douglas William Cirino, Artur Lupinetti-Cunha, Carlos Henrique Freitas, Simone Rodrigues de Freitas.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Cirino DW, Lupinetti-Cunha A, Freitas CH, de Freitas SR (2022) Do the roadkills of different mammal species respond the same way to habitat and matrix? In: Santos S, Grilo C, Shilling F, Bhardwaj M, Papp CR (Eds) Linear Infrastructure Networks with Ecological Solutions. Nature Conservation 47: 65-85. https://doi.org/10.3897/natureconservation.47.73010
|
Abstract
While road network expansion connects human settlements between themselves, it also leads to deforestation and land use changes, reducing the connectivity between natural habitat patches, and increasing roadkill risk. More than 30% of registered mammal roadkills in Brazil are concentrated in four species: Cerdocyon thous (crab-eating fox); Euphractus sexcinctus (six-banded armadillo); Tamandua tetradactyla (collared anteater) and Myrmecophaga tridactyla (giant anteater), the latter being categorized as vulnerable by IUCN redlist. Our aim was to understand how these animals’ roadkills could be related to the land use proportions on landscapes all over the Brazilian territory, and investigate if the roadkill patterns differ among species. We collected secondary data on mammal roadkills (N = 2698) from several studies in different regions of Brazil. Using MapBiomas’ data on land use and land cover, we extracted landscape composition around each roadkill sample. Through the proportion of land use and land cover in the area of influence where the roadkill occurred, we built binomial GLM models and selected the best ones by Akaike Information Criteria. For crab-eating fox and the six-banded armadillo, the best models include matrix coverage resulting in increased roadkill risk, while both anteaters’ species have a habitat and a matrix component in their best models, with an interaction between the variables. These four species seem to be roadkilled in different landscape arrangements, but in all scenarios, anthropic areas had an important influence over the models. For habitat-dependent and more sensible species, such as Tamandua tetradactyla and Myrmecophaga tridactyla, the amount of matrix influencing the roadkill risk depends on habitat availability in the landscape. It changes the strength and direction of the effect according to the proportion of natural areas in the region, while with generalist species such as Cerdocyon thous and Euphractus sexcinctus, the quantity of human-modified coverage increases the risk.
Keywords
Conservation biology, environmental impact assessment, landscape ecology, road ecology, tropics
Introduction
Road ecology is a research area that aims to understand the impact of highways and railways on natural ecosystems, economics, and society. Many studies on this subject focus on one of the most conspicuous effect of roads: wildlife mortality by vehicle collisions (
Changes in landscape composition and its structure are some of the main factors leading to biodiversity loss (
The majority of studies on road mortality focus on a small region, studying a road or a portion of it. Those studies are important to understand the local impacts of roads, and to search for patterns on specific landscape configuration. When we search for similar studies in Brazil, it is notable that some species are constantly found on roadkill registers (
The crab-eating-fox (Cerdocyon thous) is one of the most frequent species in roadkill registers according to the “Banco de Dados Brasileiro de Atropelamentos de Fauna Selvagem” – BAFS (http://cbee.ufla.br/portal/sistema_urubu/urubu_map.php) and other published researches (
The six-banded-armadillo (Euphractus sexcinctus) is also a frequent species on roadkill records (
Both anteater species (Tamandua tetradactyla and Myrmecophaga tridactyla) are more exigent in terms of habitat quality than the other two species studied in this research. They are less abundant, but also highly roadkilled. The giant-anteater (Myrmecophaga tridactyla) is a terrestrial Xernarthra that can move long distances by ground, which can aggravate the roadkill rate of this species. On the other hand, its roadkill is associated with native vegetation proximity to roads in a Cerrado area (
Most road impacts mitigation measures focus on general recommendations, such as implementation of underpasses or fencing in roadkill hotspots, which usually comes in association with native or riparian vegetation, assuming that most animals would use those areas to move and cross the road. However, we cannot assume that all species have the same habitat requirements and patterns of space usage, since it is known that the rate of underpasses usage differs among species (
Understanding the landscape patterns linked to road mortality of those species can provide guidance for protection and conservation efforts aiming to mitigate the road impacts on wildlife. Together, these four species presented here represent between 34,7% and 38,8% of the total roadkills of medium-large sized mammals in Brazil (
Methods
Roadkill data collection
We collected a sample of georeferenced roadkill data from two main sources: (1) monitoring studies across the country; and (2) the “Banco de Dados Brasileiro de Atropelamento de Fauna Selvagem” (BAFS). The first one consists of previously published systematic studies in roads of different regions of Brazil; such data was provided by collaborators (see Acknowledgements –
Landscape data and scale of analysis
For land cover and land use, we utilized the serial time data from MapBiomas (
For each species, we considered a different influence buffer radius starting from the place of the roadkill, since each one has different home ranges, body sizes, and habits requirements. We estimated the mean home range for C. thous as 4.9 km2 (
If the area of a circle is given by:
where r is the radius of the circle, so the double of a radius of a circle of a given area is:
We used this radius size because the roadkill point may have occurred on the center of the home range, or on its border (Fig.

Scheme exemplifying the radius of roadkill influence chosen. For a given roadkill point using the simple radius of home range (r), we might exclude some of the landscape characteristics if the roadkill occurred in the border of the home range. Including the possible home ranges (approximated to a circular shape), and doubling the radius (φ), we ensure that all landscape composition associated with the roadkill occurrence is incorporated within the analysis.
For each presence or absence of roadkills we calculated the proportion of land use and land cover inside the buffer based on MapBiomas land cover map for the corresponding year of the roadkill. The classes of land use and land cover considered in the analysis were: (1) forest; (2) savanna; (3) natural open areas; (4) forestry; (5) agriculture; (6) pasture; (7) farming; and (8) water. Farming represents the sum of agriculture and pasture in addition to mosaics or rotation of both classes in the same area. We conduct all landscape analysis and data extraction on ArcGIS v10.3 environment.
Statistical analysis
To estimate the relative chance of roadkill of each species we constructed binomial generalized linear models (GLM), considering the matrix of presences and absences as our response variables, and the proportion of the eight landscape variables inside the radius of roadkill influence as our predictive variables. We built four groups of models, one for each species, with one or two predictive variables by model, combining variables in pairs, and considering the interaction between them. We discarded models with some degree of correlation (> 0.6 or < -0.6) between predictive variables (see Suppl. material
All models were ranked by Akaike Information Criteria (AIC) and selected by their corrected AIC value (AICc), with lower values of AICc representing the best models (
Results
Distribution of roadkill occurrences
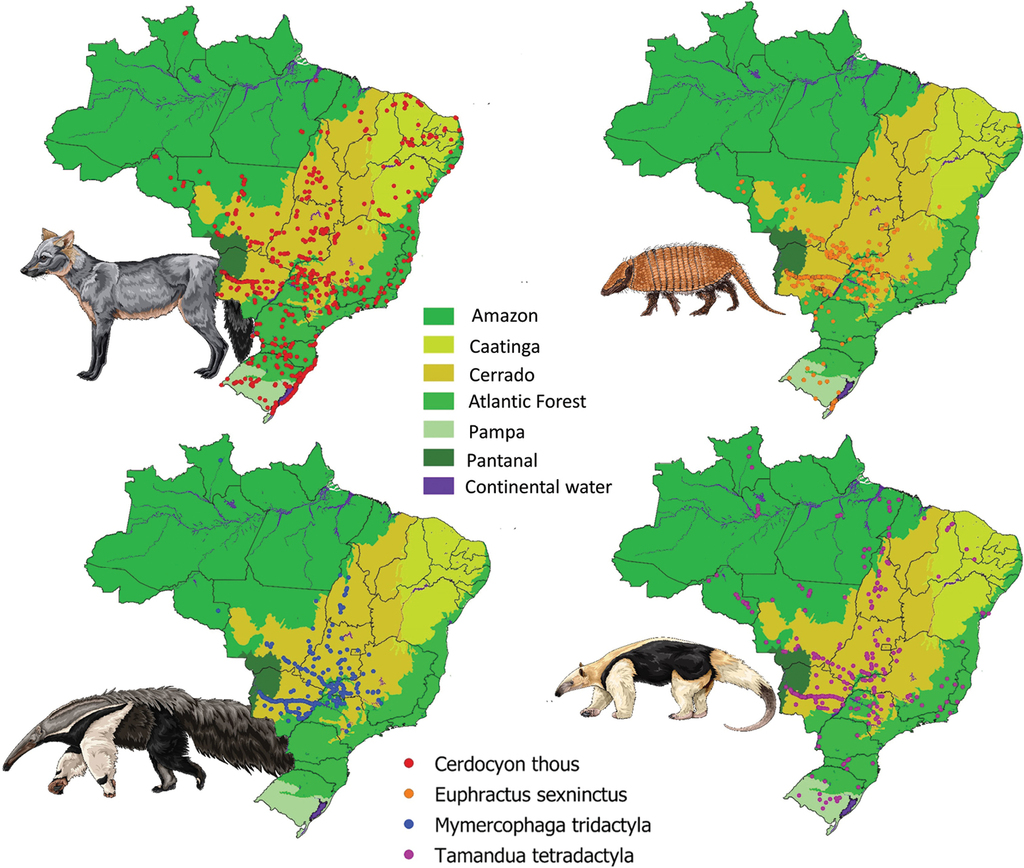
We collected a total of 2698 georeferenced roadkill records across the country (Cerdocyon thous (N = 1282); Euphractus sexcinctus (N = 589); Myrmecophaga tridactyla (N = 422) and Tamandua tetradactyla (N = 405)) (Fig.
Model selection
For Cerdocyon thous and Tamandua tetradactyla only one model was selected as the best model by AIC criteria (ΔAICc ≤ 2 and evidence ≤ 2), while the remaining studied species had two equally plausible models. For Cerdocyon thous the best model shows a positive effect of agriculture and pasture proportion inside the buffer on the chance of roadkill: for each 10% of pasture cover in landscape the roadkill risk increases by 2.7%, while for agriculture it increases by 4.6%. (Table

Best model selected by AIC for C. thous and its estimated coefficients. A model with two variables responded better to C. thous roadkill risk, being pasture and agriculture positively related to roadkill risk.
Model selected by species according to Akaike criteria. dAICc represents the AIC distance; df represents degrees of freedom; weight represents how much the model explain de variables related to all other models; evidence is the highest weight model divided by the weight of the focal model. We just considered models with evidence lower or equal to two.
| Species | Model | AICc | dAICc | df | Weight | Evidence |
|---|---|---|---|---|---|---|
| Cerdocyon thous | Pasture + Agriculture | 3545.9 | 0.0 | 3 | 0.3080 | 1.00 |
| Euphractus sexcinctus | Farming + Forestry | 1612.4 | 0.0 | 3 | 0.3772 | 1.00 |
| Pasture + Agriculture | 1612.8 | 0.3 | 3 | 0.3173 | 1.19 | |
| Myrmecophaga tridactyla | Forest + Pasture + Forest:Pasture | 1170.6 | 0.0 | 4 | 0.2057 | 1.00 |
| Forest + Savanna + Forest:Savanna | 1171.5 | 0.9 | 4 | 0.1309 | 1.57 | |
| Tamandua tetradactyla | Savanna + Agriculture + Savanna:Agriculture | 1111.4 | 0.0 | 4 | 0.8495 | 1.00 |
Discussion
Landscape features – habitat, matrix and species dependency
Cerdocyon thous – The roadkill of this species responds positively to two matrix land-uses, pasture, and agriculture in the landscape (Fig.
Besides giving information on the studied animal mortality, roadkill records are also useful for assessing a species occurrence. We found registers of C. thous roadkill occurrences out of its original geographical distribution (

Roadkill records of C. thous in Amazon ecosystem. Red dots represent roadkill records of C. thous and the hashed area is the original species’ range on Amazon. It is possible to notice some registers out of the crab-eating-fox’s range according IUCN (Lucherini, 2015), specifically in areas where forests (dark green areas) were converted into farming areas (yellow areas).
As a generalist species, Cerdocyon thous occurs, and is roadkilled in fragmented human-modified landscapes with agricultural and pasture uses. As reported for Chrysocyon brachyurus (Maned-wolf) in the Atlantic Forest (
Euphractus sexcinctus – Like C. thous, for this species two land use matrixes are included in the selected model, showing a positive relationship between farming and forestry with roadkill risk (Fig.

Best model selected by AIC for E. sexcinctus and its estimated coefficients. For the six-banded armadillo the best model represents forestry and farming positively related to roadkill risk, showing that the roadkill of this species is related to human modified landscapes.
This species inhabits a vast number of natural formations, but also human-modified landscapes, such as sugar cane plantations (
Myrmecophaga tridactyla – the best model to predict the relative risk of roadkill for this species matches with its behavior, including its relationship with pasture, forests and the interaction between these variables (Fig.

Best model selected by AIC for M. tridactyla and its estimated coefficients. The best model shows an interaction between forest and pasture, both variables positively related to roadkill risk, but the size and effect of direction changes according to the proportion of the other variable.
Depending on the amount of matrix in the landscape, the direction of the effect of habitat on roadkill risk changes. In other words, when there are small quantities of forest, the effect of pasture is positive to predict the roadkill, while when there is an increased proportion of forest in the region the effect changes, and the roadkill risk decreases with the increase of pasture areas. This could be related to the species’ habits: when the landscape is mostly composed of pastureland, the animals need to move more in search of shaded shelter. This movement decreases in frequency when there are some forested areas in the landscape, allowing the individuals to rest, and therefore, decreasing the chance of encountering a road and consequently being roadkilled. This shows the importance of maintaining habitat patches in the landscape, such as riparian forests or even native vegetation fragments inside private rural property, as established by the Brazilian Forest Code (
Tamandua tetradactyla – The roadkill risk of the collared-anteater is strongly related to savanna and agriculture patches, and its interaction (Fig.

Best model selected by AIC for T. tetradactyla and its estimated coefficients. The roadkill risk is negatively related to agriculture. However, the interaction between agriculture and savanna was significant, which changes the effect direction of agriculture in the presence of more savanna areas, increasing the risk of roadkill with the increment of agriculture, in other words, landscapes with savanna and agriculture mosaics are more likely to have collared-anteaters roadkills.
This habitat dependence reflects on the best model selected to predict the collared-anteater roadkill risk: the presence of savanna formations modulates the effect of agriculture. When a landscape has no natural formations cover, the effect of agriculture is negative, since the species probably do not occur in the area; and with the increment of habitat areas, the roadkill risk increases rapidly, reaching our model’s peak when we have at least 40% of savanna and 50% of agriculture.
On the other hand, the roadkill risk when the landscape is entirely comprised of savanna, without agriculture, is very low, and it increases very fast when there is an increase of agricultural coverage. As it is a forest dependent animal, it was unexpected that its roadkill response was better suited to savanna than to dense forests, but that can be explained by this animal’s movement pattern. In areas with continuous dense forests the locomotion of individuals occurs mainly through canopies, but in areas with low density of trees, as open areas, savannas and monocultures, it moves by ground. It can also move more often in search for sheltering trees, therefore increasing the chance of being roadkilled.
Limitations and directions for future researches
It is already known that many factors affect the roadkill risk of a species, such as species density and movement patterns (
Conclusions
For habitat dependent and more sensitive species like anteaters, the effect of the matrix on the roadkill risk depends on habitat availability in the landscape. It changes the strength and direction of the effect according to the proportion of natural areas in the region. As for generalist species, the quantity of human-modified land uses increases the roadkill risk regardless of the habitat availability or natural formations. It also indicates the occurrence of these species in those anthropic areas.
Therefore, the habitat and matrix composition impacts the studied species differently, depending on their demand and habitat dependence. Each species showed different prediction factors regarding their roadkill risk. Overall, all four target species had some dependency on the habitat, but two of them (Cerdocyon thous and Euphractus sexcinctus) are more tolerant to landscape cover changes, using some human-modified areas as habitat areas. However, the proportion and quality of natural areas should be determinant factors for Cerdocyon thous and Euphractus sexcinctus’ rate of movement, since it influences the chance of crossing a road and dying by roadkill. This movement ecology component needs to be addressed in further studies that relate the type and quality of habitat with species’ movement and roadkill rates. Currently, there is not much information regarding those common species with high roadkill rates, especially for C. thous, that can potentially cause great amounts of accidents and human injuries on Brazilian roads.
The habitat dependent species have more complex models predicting their roadkill risk, including an interaction component between habitat and matrix. It shows the importance of maintaining the natural coverage of rural properties that, as indicated by Brazilian Forest Code, can potentially decrease the risk of roadkill, connect habitat areas, and increase habitat quality. Given that, areas with vast cover of monoculture and pasture can both decrease the natural populations’ size and increase the movement of individuals that can be roadkilled while they are searching for best habitats on the landscape. Since we have shown that not only riparian corridors or continuous habitats are associated with roadkill, but also areas out of protected areas we suggest that more studies investigating the effect of movement in roadkill should be performed. We also highlight the need to consider the landscape as a whole while assessing species protection.
Acknowledgements
We thank FAPESP for the financial support (Process n° 16/12785-0) through DWC research grant, NERF/UFRGS (“Núcleo de Ecologia de Ferrovias e Rodovias” – Universidade Federal do Rio Grande do Sul) and others involved in field data collection, such as Andreas Kindel, Arnaud Desbiez, Janaina Casella and Sidnei Dornelles. We also thank Fernanda Delborgo Abra and ViaFauna for providing the illustrations in this article and contributions to data interpretation; and Guillermo Flórez for statistical contributions. We also thank the MapBiomas initiative, for the land use and land cover mapping and a reward attributed to a preliminary version of this study (CIRINO, 2018).
References
- Abra FD, Granziera BM, Huijser MP, de Barros Ferraz KMPM, Haddad CM, Paolino RM (2019) Pay or prevent? Human safety, costs to society and legal perspectives on animal-vehicle collisions in São Paulo state, Brazil. PLoS ONE 14(4): e0215152. https://doi.org/10.1371/journal.pone.0215152
- Abra FD, da Costa Canena A, Garbino GST, Medici EP (2020) Use of unfenced highway underpasses by lowland tapirs and other medium and large mammals in central-western Brazil. Perspectives in Ecology and Conservation 18(4): 247–256. https://doi.org/10.1016/j.pecon.2020.10.006
- Anacleto TCS (2007) Food Habits of Four Armadillo Species in the Cerrado Area, Mato Grosso, Brazil. Zoological Studies (Taipei, Taiwan) 46: 529–537.
- Ascensão F, Desbiez ALJ, Medici EP, Bager A, Ascensão F, Desbiez ALJ, Medici EP, Bager A (2017) Spatial patterns of road mortality of medium-large mammals in Mato Grosso do Sul, Brazil. Wildlife Research 44(2): 135–146. https://doi.org/10.1071/WR16108
- Ascensão F, Yogui D, Alves M, Medici EP, Desbiez A (2019) Predicting spatiotemporal patterns of road mortality for medium-large mammals. Journal of Environmental Management 248: e109320. https://doi.org/10.1016/j.jenvman.2019.109320
- Beisiegel BM, Lemos FG, Azevedo FC, Queirolo D, Jorge RSP (2013) Avaliação do risco de extinção do Cachorro-do-mato Cerdocyon thous (Linnaeus, 1766) no Brasil. Biodiversidade Brasileira 3: 138–145.
- Bereta A, Freitas SR, Bueno C (2017) Novas ocorrências de Chrysocyon brachyurus (Carnivora) no estado do Rio de Janeiro indicando a expansão de sua distribuição geográfica. Boletim da Sociedade Brasileira de Mastozoologia 78: 5–8. https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T4819A82316878.en
- Bergallo HG, Geise L, Bonvicino CR, Cerqueira R, D’Andrea PS, Esberard CE, Fernandez FAS, Grelle CEV, Siciliano S, Vaz SM (2000) Mamíferos. In: Bergallo HG, Rocha CFD, Van Sluys M, Geise L, Alves MA (Eds) Lista da Fauna Ameaçada do Estado do Rio de Janeiro. UERJ, Rio de Janeiro, 125–135.
- Bueno C, Faustino MT, Freitas S (2013) Influence of landscape characteristics on Capybara road-kill on highway BR-040, Southeastern Brazil. Oecologia Australis 17: 320–327. https://doi.org/10.4257/oeco.2013.1702.11
- Bueno C, Sousa COM, Freitas SR (2015) Habitat or matrix: Which is more relevant to predict road-kill of vertebrates? Brazilian Journal of Biology 75(4 suppl 1): 228–238. https://doi.org/10.1590/1519-6984.12614
- Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York, 488 pp.
- Caceres NC (2011) Biological characteristics influence mammal road kill in an Atlantic Forest-Cerrado interface in south-western Brazil. The Italian Journal of Zoology 78(3): 379–389. https://doi.org/10.1080/11250003.2011.566226
- Camilo-Alves CSP, Mourão GM (2006) Responses of a Specialized Insectivorous Mammal (Myrmecophaga tridactyla) to Variation in Ambient Temperature1. Biotropica 38: 52–56. https://doi.org/10.1111/J.1744-7429.2006.00106.X
- Carvalho CF, Custódio AEI, Junior OM (2015) Wild vertebrates roadkill aggregations on the BR-050 highway, state of Minas Gerais, Brazil. Bioscience Journal 31(3): 951–959. https://doi.org/10.14393/BJ-v31n3a2015-27468
- Castro ÉP, Bager A (2019) Sistema Urubu: A Ciência Cidadã Em Prol Da Conservação Da Biodiversidade. Revista Brasileira de Tecnologias Sociais 6(2): 111–130. https://doi.org/10.14210/rbts.v6n2.p111-130
- Cherem JJ, Kammers M, Ghizoni Jr IR, Martins A (2007) Mamíferos de médio e grande porte atropelados em rodovias do Estado de Santa Catarina, sul do Brasil. Biotemas 20: 81–96.
- Cirino DW, Freitas SR (2018) Quais são os mamíferos silvestres mais atropelados no Brasil? In: Freitas SR, Barcelos LA, Yukimitsu AH, Flórez GL (Eds) Anais do 5° Workshop de Evolução e Diversidade. Santos, 48–56. https://doi.org/978–85–65212–95–3
- Coelho IP, Kindel A, Coelho AVP (2008) Roadkills of vertebrate species on two highways through the Atlantic Forest Biosphere Reserve, southern Brazil. European Journal of Wildlife Research 54(4): 689–699. https://doi.org/10.1007/s10344-008-0197-4
- Dalponte JC, Tavares-Filho JA (2004) Diet of the Yellow Armadillo, Euphractus sexcinctus, in South-Central Brazil, 37–41. https://doi.org/10.1896/1413-4411.6.1.37
- da Silva S (2015) Impactos da duplicação de rodovias: variação da mortalidade de fauna na BR 101 Sul. Universidade Federal de São Carlos, 70 pp. http://bdtd.ibict.br/vufind/Record/SCAR_bfd36964d168c549419fa3c758aaed27
- de Santis PC (2004) Atropelamento de vertebrados silvestres em uma região fragmentada do nordeste do estado de São Paulo: quantificação do impacto e análise de fatores envolvidos. Universidade Federal de São Carlos. https://repositorio.ufscar.br/handle/ufscar/2117
- Desbiez ALJ, Medri ÍM (2010) Density and Habitat use by Giant Anteaters (Myrmecophaga tridactyla) and Southern Tamanduas (Tamandua tetradactyla) in the Pantanal Wetland, Brazil. Edentata 11: 4–10. https://doi.org/10.1896/020.011.0102
- Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B (2014) Defaunation in the Anthropocene. Science 345(6195): 401–406. https://doi.org/10.1126/science.1251817
- Dornas RAP, Kindel A, Bager A, Freitas SR (2012) Avaliação da mortalidade de vertebrados em rodovias no Brasil. In: Bager A (Ed.) Ecologia de estradas: tendências e pesquisas. Editora UFLA, Lavras, 139–152.
- Fahrig L, Rytwinski T (2009) Effects of Roads on Animal Abundance: An Empirical Review and Synthesis. Ecology and Society 14(1): 1–21. https://doi.org/10.5751/ES-02815-140121
- FATMA (2011) Lista Das Espécies Da Fauna Ameaçada De Extinção Em Santa Catarina - Relatório Técnico Final. Santa Catarina, 58 pp.
- Ferraz KMPMB, Siqueira MF, Martin PS, Esteves CF, Couto HTZ (2010) Assessment of Cerdocyon thous distribution in an agricultural mosaic, southeastern Brazil. Mammalia 74(3): 275–280. https://doi.org/10.1515/MAMM.2010.036
- Forman RTT, Alexander LE (1998) Roads and Their Major Ecological Effects 29: 207–231. http://doi.org/10.1146/annurev.ecolsys.29.1.207
- Forman RTT, Sperling D, Bissonette JA, Clevenger AP, Cutshall CD, Dale VH, Fahrig L, France RL, Goldman CR, Heanue K, Jones J, Swanson F, Turrentine T, Winter TC (2003) Road ecology: science and solutions. Island Press, Washington, 481 pp.
- Fox J, Hong J (2009) Effect displays in R for multinomial and proportional-odds logit models: Extensions to the effects package. Journal of Statistical Software 32(1): 1–24. https://doi.org/10.18637/jss.v032.i01
- Freitas CH (2009) Aleph Atropelamento de vertebrados nas rodovias MG-428 e SP-334 com análise dos fatores condicionantes e valoração econômica da fauna. Universidade Estadual Paulista (UNESP), 106 pp. https://repositorio.unesp.br/handle/11449/106548
- Freitas SR, Hawbaker TJ, Metzger JP (2010) Effects of roads, topography, and land use on forest cover dynamics in the Brazilian Atlantic Forest. Forest Ecology and Management 259(3): 410–417. https://doi.org/10.1016/j.foreco.2009.10.036
- Freitas SR, Oliveira AN, Ciocheti G, Vieira MV, Matos DMS (2014) How landscape patterns influence road-kill of three species of mammals in the brazilian savanna. Oecologia Australis 18(01): 35–45. https://doi.org/10.4257/oeco.2014.1801.02
- Freitas CH, Justino CS, Setz EZF, Freitas CH, Justino CS, Setz EZF (2015) Road-kills of the giant anteater in south-eastern Brazil: 10 years monitoring spatial and temporal determinants. Wildlife Research 41(8): 673–680. https://doi.org/10.1071/WR14220
- González-Suárez M, Ferreira FZ, Grilo C (2018) Spatial and species-level predictions of road mortality risk using trait data. Global Ecology and Biogeography 27(9): 1093–1105. https://doi.org/10.1111/geb.12769
- Grilo C, Coimbra MR, Cerqueira RC, Barbosa P, Dornas RAP, Gonçalves LO, Teixeira FZ, Coelho IP, Schmidt BR, Pacheco DLK, Schuck G, Esperando IB, Anza JA, Beduschi J, Oliveira NR, Pinheiro PF, Bager A, Secco H, Guerreiro M, Carvalho CF, Veloso AC, Custódio AEI, Marçal O, Ciocheti G, Assis J, Ribeiro MC, Francisco BSS, Cherem JJ, Trigo TC, Jardim MMA, Franceschi IC, Espinosa C, Tirelli FP, Rocha VJ, Sekiama ML, Barbosa GP, Rossi HR, Moreira TC, Cervini M, Rosa CA, Silva LG, Ferreira CMM, César A, Casella J, Mendes SL, Zina J, Bastos DFO, Souza RAT, Hartmann PA, Deffaci ACG, Mulinari J, Luzzi SC, Rezzadori T, Kolcenti C, Reis TX, Fonseca VSC, Giorgi CF, Migliorini RP, Kasper CB, Bueno C, Sobanski M, Pereira APFG, Andrade FAG, Fernandes MEB, Corrêa LLC, Nepomuceno A, Banhos A, Hannibal W, Fonseca R, Costa LA, Medici EP, Croce A, Werther K, Oliveira JP, Ribeiro JM, de Santi M, Kawanami AE, Perles L, do Couto C, Figueiró DS, Eizirik E, Correia AA, Corrêa FM, Queirolo D, Quagliatto AL, Saranholi BH, Galetti PM, Rodriguez-Castro KG, Braz VS, França FGR, Buss G, Rezini JA, Lion MB, Cheida CC, Lacerda ACR, Freitas CH, Venâncio F, Adania CH, Batisteli AF, Hegel CGZ, Mantovani JA, Rodrigues FHG, Bagatini T, Curi NHA, Emmert L, Erdmann RH, Costa RRGF, Martinelli A, Santos CVF, Kindel A (2018) BRAZIL ROAD-KILL: A data set of wildlife terrestrial vertebrate road-kills. Ecology 99(11): 2625–2625. https://doi.org/10.1002/ecy.2464
- Gumier-Costa F, Sperber CF (2009) Atropelamentos de vertebrados na Floresta Nacional de Carajás, Pará, Brasil. Acta Amazonica 39(2): 459–466. https://doi.org/10.1590/S0044-59672009000200027
- Huijser MP, Duffield JW, Clevenger AP, Ament RJ, McGowen PT (2009) Cost-Benefit Analyses of Mitigation Measures Aimed at Reducing Collisions with Large Ungulates in the United States and Canada: A Decision Support Tool. Ecology and Society 14(2): 1–26. https://doi.org/10.5751/ES-03000-140215
- Huijser MP, Abra FD, Duffield JW (2013) Mammal Road Mortality and Cost-Benefit Analyses of Mitigation Measures Aimed at Reducing Collisions with Capybara (Hydrochoerus hydrochaeris) in São Paulo State, BraziL. Oecologia Australis 17(1): 129–146. https://doi.org/10.4257/oeco.2013.1701.11
- ICMBio [Instituto Chico Mendes de Conservação da Biodiversidade] (2018) Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume I. 1st edn. Brasília.
- Jaeger JAG, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, Charbonneau N, Frank K, Gruber B, Von Toschanowitz KT (2005) Predicting when animal populations are at risk from roads: An interactive model of road avoidance behavior. Ecological Modelling 185(2–4): 329–348. https://doi.org/10.1016/j.ecolmodel.2004.12.015
- Laurance WF, Albernaz AKM, Schroth G, Fearnside PM, Bergen S, Venticinque EM, Da Costa C (2002) Predictors of deforestation in the Brazilian Amazon. Journal of Biogeography 29(5–6): 737–748. https://doi.org/10.1046/j.1365-2699.2002.00721.x
- Laurance WF, Goosem M, Laurance SGW (2009) Impacts of roads and linear clearings on tropical forests. Trends in Ecology & Evolution 24(12): 659–669. https://doi.org/10.1016/j.tree.2009.06.009
- Lemos FG, Azevedo FC, Costa HCM, Junior JAM (2011) Human threats to hoary and crab-eating foxes in central Brazil. Canid News 14: 1–6.
- Lima Santos RA, Ascensão F, Ribeiro ML, Bager A, Santos-Reis M, Aguiar LMS (2017) Assessing the consistency of hotspot and hot-moment patterns of wildlife road mortality over time. Perspectives in Ecology and Conservation 15(1): 56–60. https://doi.org/10.1016/j.pecon.2017.03.003
- Lindenmayer DB, Fischer J, Felton A, Crane M, Michael D, Macgregor C, Montague-Drake R, Manning A, Hobbs RJ (2008) Novel ecosystems resulting from landscape transformation create dilemmas for modern conservation practice. Conservation Letters 1(3): 129–135. https://doi.org/10.1111/j.1755-263X.2008.00021.x
- Lucherini M (2015) Cerdocyon thous. The IUCN Red List of Threatened Species. https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T4248A81266293.en
- Marques AAB, Fontana CS, Vélez E, Bencke GA, Schneider M, Reis RE (2002) Lista das Espécies da Ameaçadas Extinção no Rio Grande do Sul – Decreto No 41.672 de 11 de Junho de 2002. Publicações Avulsas FZB, Porto Alegre, 52 pp.
- McKinney ML, Lockwood JL (1999) Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends in Ecology & Evolution 14(11): 450–453. https://doi.org/10.1016/S0169-5347(99)01679-1
- Medri ÍM, Mourão G de M, Rodrigues FHG (2006) Ordem Xenarthra. In: Reis NR dos, Peracchi AL, Pedro WA, Lima IP de (Eds) Mamíferos do Brasil. Londrina, 71–100.
- Metzger JP (2010) O Código Florestal tem base científica? Natureza & Conservação 8(01): 1–5. https://doi.org/10.4322/natcon.00801017
- Miranda F, Bertassoni A, Abba AM (2014a) Myrmecophaga tridactyla. The IUCN Red List of Threatened Species. https://doi.org/10.2305/IUCN.UK.2014-1.RLTS.T14224A47441961.en
- Miranda F, Fallabrino A, Arteaga M, Tirira DG, Meritt DA, Superina M (2014b) Tamandua tetradactyla. The IUCN Red List of Threatened Species. https://doi.org/10.2305/IUCN.UK.2014-1.RLTS.T21350A47442916.en
- Miranda FR, Chiarello AG, Röhe F, Braga FG, Mourão G de M, de Miranda GHB, da Silva KFM, Faria-Corrêa M de A, Vaz SM, Belentani SC da S (2015) Avaliação do Risco de Extinção de Myrmecophaga tridactyla Linnaeus, 1758 no Brasil. In: Biodiversidade I-ICM de C da (Ed.) Avaliação do Risco de Extinção dos Xenartros Brasileiros. Brasília, 89–105.
- Nagendra H, Southworth J, Tucker C (2003) Accessibility as a determinant of landscape transformation in western Honduras: linking pattern and process. Landscape Ecology 2003 18(2): 141–158. https://doi.org/10.1023/A:1024430026953
- Noonan MJ, Ascensão F, Yogui DR, Desbiez ALJ (2021) Roads as ecological traps for giant anteaters. Animal Conservation. https://doi.org/10.1111/ACV.12728
- Ofstad EG, Herfindal I, Solberg EJ, Sæther B (2016) Home ranges, habitat and body mass: Simple correlates of home range size in ungulates. Proceedings of the Royal Society – Biological Sciences 283(1845): e20161234. https://doi.org/10.1098/rspb.2016.1234
- Ohana JAB, Bertassoni A, Miranda FR, Mourão G de M, de Miranda GHB, Costa JF, da Silva KFM, Faria-Corrêa M de A, Belentani SC da S (2015) Avaliação do Risco de Extinção de Tamandua tetradactyla (Linnaeus, 1758) no Brasil. In: Biodiversidade I-ICM de C da (Ed.) Avaliação do Risco de Extinção dos Xenartros Brasileiros. Brasília, 107–117.
- Passamani M, Mendes SL (2007) Espécies da Fauna Ameaçadas de Extinção no Estado do Espírito Santo. Instituto de Pesquisas da Mata Atlântica, Vitória, 140 pp.
- Pfaff A, Robalino J, Walker R, Aldrich S, Caldas M, Reis E, Perz S, Bohrer C, Arima E, Laurance W, Kirby K (2007) Road investments, spatial spillovers, and deforestation in the brazilian amazon. Journal of Regional Science 47(1): 109–123. https://doi.org/10.1111/j.1467-9787.2007.00502.x
- Pinto FAS, Clevenger AP, Grilo C (2020) Effects of roads on terrestrial vertebrate species in Latin America. Environmental Impact Assessment Review 81: e106337. https://doi.org/10.1016/j.eiar.2019.106337
- Pitman RL, Beisiegel BM (2013) Avaliação do risco de extinção do cachorro-do-mato-de-orelhas-curtas Atelocynus microtis (Sclater, 1883) no Brasil. Biodiversidade Brasileira 3: 133–137. https://www.icmbio.gov.br/portal/images/stories/biodiversidade/fauna-brasileira/avaliacao-do-risco/carnivoros/cachorro-do-mato-de-orelhas-curtas_atelocynus_microtis.pdf
- Projeto MapBiomas (2021) Coleção 3.1 da Série Anual de Mapas de Uso e Cobertura da Terra do Brasil. https://www.mapbiomas.org
- R Core Team (2020) R: A language and environment for statistical computing.
- Regolin AL, Oliveira-Santos LG, Ribeiro MC, Bailey LL (2021) Habitat quality, not habitat amount, drives mammalian habitat use in the Brazilian Pantanal. Landscape Ecology 2021(9): 1–15. https://doi.org/10.1007/s10980-021-01280-0
- Rezini JA (2010) Atropelamento de mamíferos em rodovias do leste dos Estados do Paraná e Santa Catarina, Sul do Brasil. Universidade Federal do Paraná. https://acervodigital.ufpr.br/handle/1884/23940
- Ribeiro P, Miranda JES, de Araújo DR (2017) The effect of roadkills on the persistence of xenarthran populations: The case of the Brazilian Cerrado. Edentata 18: 51–61. https://doi.org/10.2305/IUCN.CH.2017.Edentata-18-1.7.en
- Rodrigues FHG, Medri IM, Miranda G, Alves CC, Mourão G (2008) Anteater behavior and ecology. In: Vizcaíno SF, Loughry WJ (Eds) The Biology of the Xenarthra. University Press of Florida, Florida, 257–268.
- Rodrigues TF, Mantellatto AMB, Superina M, Chiarello AG (2020) Ecosystem services provided by armadillos. Biological Reviews of the Cambridge Philosophical Society 95(1): 1–21. https://doi.org/10.1111/brv.12551
- Rosa AO, Mauhs J (2004) Atropelamento de animais silvestres na rodovia rs-040. Caderno de Pesquisa série Biologia 16: 35–42. https://tspace.library.utoronto.ca/handle/1807/5369
- Rytwinski T, Fahrig L (2013) Why are some animal populations unaffected or positively affected by roads? Oecologia 173(3): 1143–1156. https://doi.org/10.1007/s00442-013-2684-x
- Sales LP, Galetti M, Pires MM (2020) Climate and land-use change will lead to a faunal “savannization” on tropical rainforests. Global Change Biology 26(12): 7036–7044. https://doi.org/10.1111/gcb.15374
- Silva KFM, Costa JF, Anacleto TCS, Timo TPC (2015) Avaliação do Risco de Extinção de Euphractus sexcinctus (Linnaeus, 1758) no Brasil. In: Biodiversidade I-ICM de C da (Ed.) Avaliação do Risco de Extinção dos Xenartros Brasileiros. Brasília, 197–207.
- Silva-Diogo O, Goebel LGA, de Sousa MR, Gusmão AC, da Costa TM, Jesus A de S, Cavalcante T (2020) Expansão da Área de Ocorrência do Lobo-Guará, Chrysocyon brachyurus (Carnivora, Canidae) No Bioma Amazônico. Oecologia Australis 24(04): e937. https://doi.org/10.4257/oeco.2020.2404.15
- Tay R (2017) Correlation, variance inflation and multicollinearity in regressionmodel. Journal of the Eastern Asia Society for Transportation Studies 12: 2006–2015. https://doi.org/10.11175/easts.12.2006
- Teixeira FZ, Coelho IP, Esperandio IB, Oliveira NR, Peter FP, Dornelles SS, Delazeri NR, Tavares M, Martins B, Kindel A (2013) Are road-kill hotspots coincident among different vertebrate groups? Oecologia Australis 17(1): 36–47. https://doi.org/10.4257/oeco.2013.1701.04
- Teixeira FZ, Kindel A, Hartz SM, Mitchell S, Fahrig L (2017) When road-kill hotspots do not indicate the best sites for road-kill mitigation. Journal of Applied Ecology 54(5): 1544–1551. https://doi.org/10.1111/1365-2664.12870
- Teixeira FZ, Rytwinski T, Fahrig L (2020) Inference in road ecology research: What we know versus what we think we know. Biology Letters 16(7): e20200140. https://doi.org/10.1098/rsbl.2020.0140
- Turci LCB, Bernarde PS (2009) Vertebrados atropelados na Rodovia Estadual 383 em Rondônia, Brasil. Biotemas 22(1): 121–127. https://doi.org/10.5007/2175-7925.2009v22n1p121
- Van Der Ree R, Smith DJ, Grilo C (2015) Handbook of Road Ecology Handbook of Road Ecology. John Wiley & Sons, Chichester, 522 pp. https://doi.org/10.1002/9781118568170
- Versiani NF, Bailey LL, Pasqualotto N, Rodrigues TF, Paolino RM, Alberici V, Chiarello AG (2021) Protected areas and unpaved roads mediate habitat use of the giant anteater in anthropogenic landscapes. Journal of Mammalogy 102(3): 802–813. https://doi.org/10.1093/jmammal/gyab004
- Viana DS, Granados JE, Fandos P, Pérez JM, Cano-Manuel FJ, Burón D, Fandos G, Aguado MAP, Figuerola J, Soriguer RC (2018) Linking seasonal home range size with habitat selection and movement in a mountain ungulate. Movement Ecology 6(1): e1. https://doi.org/10.1186/s40462-017-0119-8
- Vieira EM (1996) Highway mortality of mammals in central Brazil. Ciencia e Cultura 48: 270–272.
- Wei T, Simko V (2021) . R package ‘corrplot’: Visualization of a Correlation Matrix. (Version 0.92). https://github.com/taiyun/corrplot
- Wickham H (2016) ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York. https://doi.org/10.1007/978-3-319-24277-4
Supplementary material
Correlation plot and R script for building and selecting best models
Data type: R script (text file)
Explanation note: Plot of correlations between predictor variables. The script used for reading variables, building statistical models and selecting the best model by Akaike Information Criteria.