Research Article |
|
Corresponding author: Jihong Huang ( northalluvion@163.com ) Academic editor: Chris Margules
© 2018 Jihong Huang, Canran Liu, Zhongjun Guo, Keping Ma, Runguo Zang, Yi Ding, Xinghui Lu, Jiping Wang, Ruoyun Yu.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Huang J, Liu C, Guo Z, Ma K, Zang R, Ding Y, Lu X, Wang J, Yu R (2018) Seed plant features, distribution patterns, diversity hotspots, and conservation gaps in Xinjiang, China. Nature Conservation 27: 1-15. https://doi.org/10.3897/natureconservation.27.23728
|
Abstract
The flora in Xinjiang is unique. Decisions about biodiversity conservation and management based on seed plant diversity hotspots and conservation gaps in Xinjiang are essential to maintain this unique flora. Based on a species distribution dataset of seed plants, we measured seed plant diversity using species richness and phylogenetic diversity indices. Five percent of Xinjiang’s total land area with the highest biodiversity was used to identify hotspots for each index. In total, eight hotspots were identified. Most hotspots were located in mountainous areas, mainly in the Tianshan Mountains and Altai Mountains. Furthermore, we detected conservation gaps for Xinjiang’s seed flora hotspots by overlaying nature reserve maps on to maps of identified hotspots and we designated priority conservation gaps for hotspots by overlaying global biodiversity hotspot maps on to hotspot conservation gaps maps. Most of Xinjiang’s seed plant hotspots are poorly protected; only 10.45% of these hotspots were covered by nature reserves. We suggest that it is essential to promote network function of nature reserves within these hotspots in Xinjiang to conserve this unique flora.
Keywords
plant diversity, diversity hotspots, phylogenetic diversity, conservation gaps, Xinjiang
Introduction
Biodiversity is unevenly distributed; therefore, prioritisation is essential to minimise biodiversity loss (
Xinjiang Uygur Autonomous Region (Xinjiang hereafter) is a large (1.64 million km2), topographically varied inland province in northwest China that extends from high mountains (8611 m above sea level) to lowland basins (156 m below sea level) and humid forest to grasslands and dry desert. Consequently, this region has an extreme arid and cold climate and exhibits striking climate, soil and vegetation patterns. Species richness patterns in Xinjiang were previously reported for woody plants (
In this paper, based on native seed plant species in Xinjiang, we identified seed plant hotspots using both species richness and phylogenetic diversity indices. Moreover, Xinjiang’s seed plants conservation gaps, which are hotspot areas that are not covered by China’s nature reserves, were detected by overlaying the seed plant hotspot map on a map of Xinjiang’s nature reserves; and Xinjiang seed plant priority conservation gaps, which are areas within both Xinjiang seed plant hotspots and global biodiversity hotspots that are not covered by Xinjiang’s nature reserves, were detected by overlaying the Xinjiang seed plant conservation gap map on a global biodiversity hotspot map. Finally, we discussed the potential causes of the patterns revealed in this study and proposed suggestions for Xinjiang flora protection.
Methods
Data sets
We compiled a species distribution database that included all seed plant species distributed in Xinjiang based on a large number of herbarium specimens (http://www.cvh.org.cn/), a list of Chinese seed plant species and distribution information at provincial level (
A georeferenced or spatially referenced dataset of 33 nature reserves in Xinjiang was constructed. The main body of nature reserves was downloaded from the World Database on Protected Areas (http://www.protectedplanet.net/). The global biodiversity hotspots dataset was downloaded from Conservation International (http://www.conservation.org/How/Pages/Hotspots.aspx).
Diversity measures and phylogenetic tree construction
To detect the distribution patterns of Xinjiang seed flora, we calculated seed species richness and phylogenetic diversity of seed plants. Phylogenetic diversity is a simple measure of evolutionary diversity that is determined based on the sum of the lengths of all branches that are members of the corresponding minimum spanning path (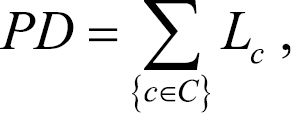
where C indicates the set of branches in the minimum spanning path joining the species to the root of the tree, c indicates a branch in the spanning path, and indicates the length of branch c. Phylogenetic diversities were calculated by using the phylogenetic diversity algorithm in PHYLOCOM (
PHYLOMATIC (
Hotspot identification and ranking and conservation gap analysis
Both diversity indices were calculated using two packages, Ape and Picante, in R 3.3.2 (
We then identified ‘conservation gaps’ (
Results
Xinjiang seed plant features
Based on our data, Xinjiang has 3715 seed plant species (including 66 subspecies and 224 varieties), which account for 12.71% of all seed plant species in China. These seed plants belong to 767 genera and 113 families. Of these species, gymnosperms, dicots and monocots accounted for 1.10%, 80.57% and 18.33%, respectively, of all Xinjiang seed plant species (Table
Xinjiang seed plants distribution and similarity at the regional level
Species richness at the regional scale generally increased with decreasing latitude in Xinjiang. In the 10 geographical regions (Fig.

Map showing the 10 geographical regions of Xinjiang. Black number identifies geographical regions: 1 North of Junggar Basin 2 South of Junggar Basin 3 Ily Valley 4 Eastern zone of Xinjiang 5 West-North Tarim Basin 6 East-North Tarim Basin 7 West Tarim Basin 8 West-Central Tarim Basin 9 South-Central Tarim Basin 10 East-Central Tarim Basin.

Species richness distribution for 3715 species in Xinjiang across the 10 spatial range classes. Spatial range was measured based on the number of geographical regions, without specific reference to particular regions. The line represents the least-squares regression using a hyperbola model with 61.07 and 1060.20 as the a and b parameters, respectively (R2=0.89, P<0.001).
Numbers of seed plant orders, families, genera and species in each geographical region of Xinjiang. Regional division of Xinjiang is shown in Fig.
| Geographical regions | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | Overall | |
| No. of orders | 40 | 43 | 42 | 38 | 36 | 39 | 38 | 35 | 26 | 25 | 45 |
| No. of families | 92 | 104 | 95 | 76 | 72 | 79 | 83 | 64 | 42 | 40 | 113 |
| No. of genera | 572 | 629 | 569 | 365 | 315 | 378 | 403 | 251 | 130 | 153 | 767 |
| No. of species | 2291 | 2445 | 2110 | 1102 | 824 | 1163 | 1398 | 618 | 264 | 343 | 3716 |
| Gymnosperms | 22 | 30 | 25 | 17 | 12 | 17 | 18 | 9 | 2 | 2 | 41 |
| Dicots | 1820 | 1963 | 1708 | 870 | 630 | 881 | 1081 | 472 | 198 | 235 | 2994 |
| Monocots | 449 | 452 | 377 | 215 | 182 | 265 | 299 | 137 | 64 | 106 | 681 |
The similarity indices for pairwise comparisons of regional species composition ranged from 0.07 to 0.58 (Table
Jaccard similarity indices for pairwise comparisons of geographical regions using all Xinjiang seed plant species. Regional division of Xinjiang is shown in Fig.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | ||||||||||
| 2 | 0.58 | |||||||||
| 3 | 0.55 | 0.58 | ||||||||
| 4 | 0.35 | 0.38 | 0.34 | |||||||
| 5 | 0.26 | 0.28 | 0.28 | 0.36 | ||||||
| 6 | 0.36 | 0.38 | 0.38 | 0.41 | 0.38 | |||||
| 7 | 0.33 | 0.36 | 0.34 | 0.35 | 0.38 | 0.40 | ||||
| 8 | 0.15 | 0.18 | 0.17 | 0.25 | 0.30 | 0.27 | 0.33 | |||
| 9 | 0.07 | 0.08 | 0.07 | 0.13 | 0.17 | 0.14 | 0.15 | 0.27 | ||
| 10 | 0.09 | 0.09 | 0.08 | 0.14 | 0.17 | 0.15 | 0.18 | 0.26 | 0.30 |
Diversity hotspots of Xinjiang seed plants
Distributions of Xinjiang seed plant species richness and phylogenetic diversity were uneven and concentrated in the northern regions of Xinjiang (i.e. north of the Tianshan Mountains; Fig.

Geographic distribution of Xinjiang seed plant diversity patterns and hotspots and major mountains ranges in northwest China. a Geographic distribution of species richness b geographic distribution of phylogenetic diversity c major mountains ranges d diversity hotspots identified by species richness e diversity hotspots identified by phylogenetic diversity and f diversity hotspots identified by both species richness and phylogenetic diversity. Albers projection. Hotspot centre codes are consistent with those listed in Table
| Code | Plant diversity hotspots | Number of species | Area of hotspots (km2) | Proportion of area uncovered with nature reserves (%) | Proportion of area covered with global biodiversity hotspots (%) | Conservation rank |
|---|---|---|---|---|---|---|
| H1 | Mountain areas in and around Altay city-Burqin County | 1406 | 32500 | 94.21 | 0.00 | 2 |
| H2 | Mountainous areas in and around Urumqi city | 1627 | 30000 | 88.5 | 63.06 | 3 |
| H3 | Mountainous areas in and around Yining city-Gongliu county | 1084 | 15000 | 98.48 | 99.29 | 1 |
| H4 | Fuyun-Qinghe-Qitai-Jimsar Counties boundaries | 969 | 10000 | 47.66 | 0.00 | 7 |
| H5 | Shihezi City and Shawan county | 751 | 7500 | 100.00 | 42.39 | 4 |
| H6 | Tacheng City and Yumin county and Ermin county boundary | 618 | 5000 | 100.00 | 0.00 | 5 |
| H7 | Bole City-Huocheng County and Wenquan County boundary | 698 | 5000 | 94.68 | 99.08 | 6 |
| H8 | Karamay City-Hoboksar County-Shawan County boundaries | 481 | 2500 | 92.19 | 0.00 | 8 |
Priority conservation gap areas for Xinjiang seed plant hotspots
The spatial distributions of 33 nature reserves and eight hotspots of Xinjiang seed plant species significantly differed (Fig.
Four of the eight hotspots overlapped with global biodiversity hotspots (H2, H3, H5 and H7; 60000.00 km2); these overlapping areas accounted for 55.81% of the total area of all hotspots and contained 59.11% (2192 species) of all Xinjiang seed plant species. The remaining four hotspots harboured 1836 seed plant species, which accounted for 49.42% of all Xinjiang seed plant species. A priority conservation area that could alleviate Xinjiang seed plant conservation gaps is centre H5 (Fig.

Discussion
Xinjiang flora uniqueness and complexity
Xinjiang is located in the contact area of several natural geographical units, including the Altai Mountains, Tianshan Mountains, Pamirs, Kunlun Mountains, Altun Mountains and Northern Tibetan Plateau. Moreover, phytogeographically, Xinjiang also has intersections of Eurasian Forest, Eurasian steppe, Central Asian desert and Himalayan Chinese plant sub-regions. These characteristics have led to floral uniqueness and vegetation complexity in Xinjiang (
Xinjiang seed plant hotspots
Species richness is higher in Xinjiang mountains than basins (
Hotspots in major mountain ranges of Xinjiang
Seed plant species are unevenly distributed across Xinjiang; all their hotspots are located in mountain areas mainly within the Tianshan Mountains, Altai Mountains and western mountainous areas in Junggar. These areas approximately correspond to the montane forest and cold temperate coniferous forest in temperate steppe and temperate desert regions (
Suggestions for Xinjiang flora protection
Xinjiang is the largest province in China and accounts for one-sixth of China’s total areas (
Firstly, we need to create a unified conservation plan for the Xinjiang flora. Conservation biologists have proposed many conservation suggestions and schemes across China (
Secondly, we need to establish a provincal georeferenced (spatially referenced) database for biodiversity that includes plant, animal and nature reserve distributions. In particular, efficient conservation of Xinjiang’s plant resources requires accurate data on the current distribution and threat status of plant species in the country (
Conclusions
Different diversity indices revealed similar distribution patterns of plant diversity in Xinjiang Province. A total of eight seed plant hotspots in Xinjiang were identified by species richness and phylogenetic diversity. All of these hotspots are located in the mountainous areas within the Tianshan Mountains and Altai Mountains. Most centres of Xinjiang seed plant hotspots are not covered by the current nature reserve system.
Acknowledgements
We would like to thank Dr. Chen Bin at the Shanghai Chenshan Botanical Garden, Dr. Qiong Ding at the Hainan University, Dr. Zhen Zhang at the Anhui Agricultural University, Dr. Zhidong Zhang at the Hebei Agricultural University for their assistance with data collection. We are particularly grateful to the associate editor and anonymous reviewers for their valuable comments on the manuscript. We thank Mallory Eckstut, PhD, from Liwen Bianji, Edanz Editing China (www.liwenbianji.cn/ac), for editing the English text of a draft of this manuscript. The research was supported by the National Nonprofit Institute Research Grant of the Chinese Academy of Forestry (CAFYBB2014MA005), the National Natural Science Foundation of China (41471048) and the National Nonprofit Institute Research Grant of the Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry (CAFIFEEP2015B02).
References
- Axelrod DI, Al-Shehbaz I, Raven PH (1996) History of the modern flora of China. In: Zhang AL, Wu SG (Eds) Floristic Characteristics and Diversity of East Asian Plants.China Higher Education Press, Beijing, 43–55.
- Brooks TM, Mittermeier RA, da Fonseca GAB, Gerlach J, Hoffmann M, Lamoreux JF, Mittermeier CG, Pilgrim JD, Rodrigues ASL (2006) Global biodiversity conservation priorities. Science 313(5783): 58–61. https://doi.org/10.1126/science.1127609
- Cadotte MW, Jonathan Davies T, Regetz J, Kembel SW, Cleland E, Oakley TH (2010) Phylogenetic diversity metrics for ecological communities: Integrating species richness, abundance and evolutionary history. Ecology Letters 13(1): 96–105. https://doi.org/10.1111/j.1461-0248.2009.01405.x
- Chen L (2014) Flora and Vegetational Geography of China. Science Press, Beijing.
- CRFX (1992–2011) Flora Xinjiangensis. Xinjiang Science & Technology & Hygiene Publishing House, Urumqi.
- Editorial Committee of China National Biodiversity Conservation Strategy and Action Plan (2011) China National Biodiversity Conservation Strategy and Action Plan. China Environmental Science Press, Beijing
- Editorial Committee of Flora of Xinjiang (1992) Flora of Xinjiang, Vol. 1. Xinjiang Science, Technology and Sanitation Press, Urumqi.
- Editorial Committee of Vegetation Map China (2008) Vegetation Map of China (1:1000 000). Geology Press, Beijing.
- Faith DP (1992) Conservation Evaluation and Phylogenetic Diversity Biol Conserv 61: 1–10.
- Fang J, Wang Z, Tao Z (2009) Atlas of Woody Plants in China:Distribution and Climate. Higher Education Press, Beijing.
- Forest F, Grenyer R, Rouget M, Davies TJ, Cowling RM, Faith DP, Balmford A, Manning JC, Procheş Ş, van der Bank M, Reeves G, Hedderson TAJ, Savolainen V (2007) Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 445(7129): 757–760. https://doi.org/10.1038/nature05587
- Huang J, Huang J, Liu C, Zhang J, Lu X, Ma K (2016a) Diversity hotspots and conservation gaps for the Chinese endemic seed flora. Biological Conservation 198: 104–112. https://doi.org/10.1016/j.biocon.2016.04.007
- Huang J, Huang J, Lu X, Ma K (2016b) Diversity distribution patterns of Chinese endemic seed plant species and their implications for conservation planning. Scientific Reports 6(1): 33913. https://doi.org/10.1038/srep33913
- Huang J, Lu X, Huang J, Ma K (2016c) Conservation priority of endemic Chinese flora at family and genus levels. Biodiversity and Conservation 25(1): 23–35. https://doi.org/10.1007/s10531-015-1027-0
- Huang J, Ma K, Chen B (2014) Diversity and Geographic Distribution of Endemic Species of Seed Plants in China. Higher Education Press, Beijing.
- Latham RE, Ricklefs RE (1993) Continental comparisons of temperate-zone tree species diversity. In: Ricklefs RE, Schluter D (Eds) Species Diversity in Ecological Communities: Historical and Geographical Perspectives.University of Chicago Press, Chicago, 294–314.
- Li L, Wang Z, Zerbe S, Abdusalih N, Tang Z, Ma M, Yin L, Mohammat A, Han W, Fang J (2013) Species Richness Patterns and Water-Energy Dynamics in the Drylands of Northwest China Plos One 8: e66450. https://doi.org/10.1371/journal.pone.0066450
- Li W, Wang X, Wang S (1993) Conservation stragety for biodiversity of China. In: Chen L, Wang X, Wang S (Eds) Biodiversity in China: Status and Conservation Strategy.Science Press, Beijing, 164–205.
- Liu J, Ouyang Z, Pimm SL, Raven PH, Wang X, Miao H, Han N (2003) Protecting China’s biodiversity. Science 300(5623): 1240–1241. https://doi.org/10.1126/science.1078868
- Lomolino MV, Riddle BR, Brown JH (2006) Biogeography (3rd edn). Sinauer Associates, Inc., Sunderland, Massachusetts.
- Lu Q, Wang J, Chu J (2012) Desert Plants in China. China Forestry Publishing House, Beijing.
- Ministry of Environmental Protection the People’s Republic of China (2015) The 2014 Report on the State of Environment in China. Beijing.
- Mittermeier RA, Gil PR, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, da Fonseca GAB, Seligmann PA, Ford H (2005) Hotspots Revisited: Earth's Biologically Richest and Most Endangered Terrestrial Ecoregions. The University of Chicago Press, Chicago.
- Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403(6772): 853–858. https://doi.org/10.1038/35002501
- Olson DM, Dinerstein E (1998) The global 200: A representation approach to conserving the Earth’s most biologically valuable ecoregions. Conservation Biology 12(3): 502–515. https://doi.org/10.1046/j.1523-1739.1998.012003502.x
- Omland KE, Cook LG, Crisp MD (2008) Tree thinking for all biology: The problem with reading phylogenies as ladders of progress. BioEssays 30(9): 854–867. https://doi.org/10.1002/bies.20794
- Orme CDL, Davies RG, Burgess M, Eigenbrod F, Pickup N, Olson VA, Webster AJ, Ding T-S, Rasmussen PC, Ridgely RS, Stattersfield AJ, Bennett PM, Blackburn TM, Gaston KJ, Owens IPF (2005) Global hotspots of species richness are not congruent with endemism or threat. Nature 436(7053): 1016–1019. https://doi.org/10.1038/nature03850
- Prendergast JR, Quinn RM, Lawton JH (1999) The gaps between theory and practice in selecting nature reserves. Conservation Biology 13(3): 484–492. https://doi.org/10.1046/j.1523-1739.1999.97428.x
- Prendergast JR, Quinn RM, Lawton JH, Eversham BC, Gibbons DW (1993) Rare Species, the Coincidence of Diversity Hotspots and Conservation Strategies Nature 365: 335–337.
- Qian H, Ricklefs RE (1999) A comparison of the taxonomic richness of vascular plants in China and the United States Am Nat 154:160-181.
- R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
- Reid WV (1998) Biodiversity hotspots. Trends in Ecology & Evolution 13(7): 275–280. https://doi.org/10.1016/S0169-5347(98)01363-9
- Rosauer DF, Laffan SW (2008) Linking phylogenetic trees, taxonomy & geography to map phylogeography using Biodiverse. Taxonomic Data Working Group 2008, Perth, Australia.
- Rosenzweig ML (1995) Species diversity in space and time. Cambridge University Press, Cambridge.
- Sang W, Ma K, Axmacher JC (2011) Securing a future for China’s wild plant resources. BioScience 61: 720–725. https://doi.org/10.1525/bio.2011.61.9.11
- Vane-Wright RI, Humphries CJ, Williams PH (1991) What to protect? Systematics and the agony of choice. Biological Conservation 55(3): 235–254. https://doi.org/10.1016/0006-3207(91)90030-D
- Wang X, Feng Z, Fu L, Wu P, Xing G (1993) Key taxa and biodiversity areas in China. In: Chen L, Wang X, Wang S (Eds) Biodiversity in China: status and conservation strategy.Science Press, Beijing, 164–205.
- Webb CO, Ackerly DD, Kembel SW (2008) Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics (Oxford, England) 2418): 2098–2100. https://doi.org/10.1093/bioinformatics/btn358
- Webb CO, Donoghue MJ (2005) Phylomatic: Tree assembly for applied phylogenetics. Molecular Ecology Notes 5(1): 181–183. https://doi.org/10.1111/j.1471-8286.2004.00829.x
- Wikstrom N, Savolainen V, Chase MW (2001) Evolution of the angiosperms: Calibrating the family tree. Proceedings. Biological Sciences 268(1482): 2211–2220. https://doi.org/10.1098/rspb.2001.1782
- Wu R, Zhang S, Yu DW, Zhao P, Li X, Wang L, Yu Q, Ma J, Chen A, Long Y (2011) Effectiveness of China’s nature reserves in representing ecological diversity. Frontiers in Ecology and the Environment 9(7): 383–389. https://doi.org/10.1890/100093
- Wu Z (1980) The Vegetation of China. Science Press, Beijing.
- Wu Z, Raven PH, Hong D (1994–2012) Flora of China. Science Press or Missouri Botanical Garden Press, Beijing or St. Louis.
- Wu Z, Wang H (1983) Physical Geography of China: phytogeography (I). Science Press, Beijing.
- Xinjiang Investigation Group of Chinese Academy of Sciences (1978) The vegetation of Xinjiang and its application. Science Press, Beijing.
- Xu Y, Shen Z, Ying L, Wang Z, Huang J, Zang R, Jiang Y (2017) Hotspot analyses indicate significant conservation gaps for evergreen broadleaved woody plants in China. Scientific Reports 7(1): 1859. https://doi.org/10.1038/s41598-017-02098-0
- Yang C (2012) Sylva Xinjiangensis. China Forestry Publishing House, Beijing.
- Yin L (2006) Rare Endangered Endemic Higher Plants in Xinjiang of China. Xinjiang Science & Technology Publishing House, Urumqi.
- Zhang H-X, Zhang M-L (2014) Insight into distribution patterns and conservation planning in relation to woody species diversity in Xinjiang, arid northwestern China. Biological Conservation 177: 165–173. https://doi.org/10.1016/j.biocon.2014.07.005
- Zhang Y, Ma K (2008) Geographic distribution patterns and status assessment of threatened plants in China. Biodiversity and Conservation 17(7): 1783–1798. https://doi.org/10.1007/s10531-008-9384-6